
Amber types
Two types of amber are found in the Hollín Formation (Fig. 3). The more common type formed around the roots of resin-producing plants (Fig. 3a–c), while the less frequent aerial amber formed from resin exposed to air (Fig. 3d–h). Both types are often translucent and yellowish, but can also appear greenish or reddish. Aerial amber pieces are usually small (<7 cm) and stalactitic in shape, whereas root-derived amber can be up to 40 cm and is typically subspherical or kidney-shaped. Due to its formation in confined conditions, the root-derived amber lacks arthropod or plant inclusions, as far as we observed, unlike aerial one (Fig. 4 and Supplementary Fig. 4).
Fig. 3: Two types of amber from the Genoveva quarry.
a–c Large amber pieces (kidney-shaped), formed from resin exuded from roots in confined conditions, are the most common pieces in the outcrop, but they lack bioinclusions. d–h Small or medium-sized amber pieces (stalactite-shaped), formed from resin exuded from trunks or branches in aerial conditions, are scarce, but some of them contain bioinclusions. The pieces (d–h) will be housed at the Laboratory of Palaeontology, Escuela Politécnica Nacional, in Quito (Ecuador). Scale bars: 1 cm (d–f), 5 mm (g, h).
Fig. 4: Insect inclusions found in the Genoveva amber.
a Diptera: Ceratopogonidae, Archiaustroconops (N-113-JR-2022-19). b Diptera: Chironomidae (N-113-JR-2022-02). c Diptera: Dolichopodidae, Microphorites n. sp. (N-113-JR-2022-13). d Diptera: Chironomidae (N-113-JR-2022-12). e Trichoptera (N-113-JR-2022-18), incomplete specimen showing the fore wing venation in this view. f, g Coleoptera: Tetratomidae (N-113-JR-2022-01); 3D virtual extraction (synchrotron) (f), optical image (g). h Hymenoptera: †Stigmaphronidae (N-113-JR-2022-04). i Hymenoptera: Scelionidae (N-113-JR-2022-08). All will be housed at the Laboratory of Palaeontology, Escuela Politécnica Nacional, Quito (Ecuador). Scale bars: 1 mm (e), 0.5 mm (a–d, h, i), 0.2 mm (f, g).
The Fourier transform infrared spectroscopy (FTIR)
The FTIR characteristics of three samples of the amber from the Genoveva quarry are compared with FTIR characteristics of the amber found in Pungarayacu and Río Misahuallí quarries (Supplementary Fig. 5), and of the amber from other Cretaceous localities (Supplementary Fig. 6): Jezzine (Barremian, Lebanon), El Soplao and San Just (Albian, Spain), Archingeray-Les Nouillers (Albian–Cenomanian, France), Kachin (Cenomanian, Myanmar) and Raritan-New Jersey (Turonian USA).
Cretaceous ambers from the three Ecuadorian localities (Genoveva, Pungarayacu and Río Misuhualli quarries) exhibits similar features (Supplementary Fig. 5), including absorption bands between 2930 and 2860 cm−1, attributed to the stretching vibrations of saturated C–H bonds, and corresponding absorption bands at 1470–1443 cm−1 and 1384–1370 cm−1, linked to CH2—CH3 bending vibration. The FTIR spectra of amber from the Genoveva quarry were analysed across three wavenumbers regions diagnostic for amber: 3000–2800 cm1/1460–1370 cm1, 1760–1680 cm1, and 1250–1000 cm1. These regions correspond to the presence of alkane, carbonyl, and ether groups, respectively. The Genoveva amber exhibits a marked increase in absorbance intensity at 1719 cm1, attributed to ester-group vibration, along with a slight increase in absorbance at 1032 cm1, 1144 cm1, and 1696 cm1, corresponding to C–O–C––C, C–O–C, and C––O of carboxylic acids.
In contrast, a slight decrease in absorbance was observed at 963 cm1, which is attributed to C––C stretching vibration. All samples displayed a small peak at 3076 cm−1, attributed to the asymmetric C–H stretching of monoalkyl groups, as well as a more prominent peak at around 2935 cm−1, representing a doublet produced by methylene groups. In addition, two smaller peaks were observed off the shoulder at 2866 cm−1 and 2844 cm−1, which correspond to the most intense group bands characteristic of all Cretaceous amber spectra. These three peaks result from the aliphatic stretching of single C–H bonds. All samples shared a peak at 1693 cm−1, associated with C = O bonds in the carboxyl groups of resin acids. At 1460 cm−1, a small trough on the shoulder of the peak at 1448 cm−1 was observed, corresponding to C–H bending motions of methyl and methylene functional groups. The peaks between 1300 and 1100 cm−1 were assignable to C–O bonds, with a prominent feature in the 1144–1226 cm−1 range. It is well established that the C = O group acts as a strong chromophore, and the colour of amber tends to darken upon oxidation due to the increased concentration of C = O groups. All samples of the Genoveva quarry also shared peaks at 1156 cm−1 and 1093 cm−1, along with additional peaks at 1032 cm−1 and around 1010 cm−1. A series of moderate to weak absorption bands within the 1240–1150 cm−1 spectral region suggests the presence of oxygen-bearing functional groups such as esters, alcohols, and ethers. This sequence of spectral peaks is linked to the C–O stretching vibration of these oxygen-containing functional groups. The absence of exocyclic methylenic bands at 880 cm−1, 1640 cm−1, and 3070 cm−1 indicates the high maturity of the Genoveva amber.
The FTIR analyses of the amber samples from the Genoveva quarry (Supplementary Fig. 5) reveal highly consistent spectra across all specimens, closely matching those of amber produced by Araucariaceae (Supplementary Fig. 6). These results suggest that the fossil resin fossilised was exuded by araucariacean conifers.
Amber geochemistry
Extractable organic matter comprises 15% of the total amber weight in the examined Genoveva samples (rock samples from amber-containing levels G2 and M8, Supplementary Fig. 3) an unusually low value compared to other Cretaceous ambers. This is consistent with the presence of a petroleum-like mixture of hydrocarbons, including the homologous series of n-alkanes from C12 to C35, pristane, phytane and other unsaturated hydrocarbons (Supplementary Fig. 7a, b). These hydrocarbons permeated the amber and altered its original composition. Despite this unusual feature, the Cretaceous amber biomarkers amberene I and 1-methylamberene II17,18 are preserved in the Genoveva amber. These biomarkers are possibly derived from the diagenesis of labdane diterpenoids. The absence of phenolic diterpenoids and triterpenes suggests no contribution from Cupressaceae or angiosperm resin-producing taxa17,19.
The presence of paraffins in the Genoveva amber is an uncommon feature that has been previously reported only in the Cretaceous amber from Araripe (Brazil)20. In the amber samples of Genoveva quarry, the hydrocarbon composition is consistent with the abundant bitumen present in the Hollín Formation rocks. The extraction of bitumen of host rock shows a 4–6% of extractable organic matter, showing a similar Carbon Preference Index (CPI) between 1 and 1.05, and pristane/phytane indexes to those observed in the amber total extract (Supplementary Fig. 7a). The n-alkane distribution trapped in amber shows a relative enrichment on C > 27 compared to those of surrounding bitumen. The host rock bitumen shows a strong depletion in light hydrocarbons, being enriched in heavy (C > 35) n-paraffins and heavy aromatic hydrocarbons. The loss of lighter components, and the n-C17/pristane and n-C18/phytane<1, suggests a high degradation/weathering of the original petroleum.
Petroleum hydrocarbons likely penetrated the amber structure and altered its overall molecular composition due to the permeability and apolar character of the resin. The composition was depleted in saturated and light resin-derived hydrocarbons, likely extracted by light petroleum components and subsequently lost along with the lighter fractions of petroleum. This process left behind an unusual distribution of linear hydrocarbons trapped in amber, and heavy bitumen, observed in analysed samples of amber-bearing rock. The low n-C17/pristane and n-C18/phytane ratios in the amber-trapped hydrocarbons are consistent with the petroleum alteration and degradation. Amberene I and 1-methylamberene II have been identified; they are derived from the diagenesis of labdane diterpenoids and also the related 2,5-dimethyl-1-(3’-methylbutyl)naphthalene III, the aromatised derivative of I, as well as the tetralins IV–V. The relative proportion of II is higher than expected for ambers, consistent with the loss of lighter amberene after petroleum-induced alteration of the amber composition. Finding traces of amberene in the surrounding bitumen is consistent with this process. The alteration induced by petroleum affected acid terpenes in lesser grade, due to its higher polarity. Consequently, pimaric acid series VI and labdanoic acids VII–IX, have been identified. The terpenoids of the abietane class are present in lower proportion and are highly aromatised, with identifiable simonellite X, which are also present in the surrounding bitumen. Analysis of polar fraction (Supplementary Fig. 7c) shows the presence of TMS derivatives of agathic acid and its isomers XI, and confirms agatholic acid XII and resinic acids of the pimarane and labdane series. The predominance of labdanoids and the similarity of the biomarker assemblage to those of Early Cretaceous amber from eastern Iberia21 also suggest that the Araucariaceae family is likely the botanical source of the Genoveva amber in the Genoveva quarry.
Arthropod inclusions
Twenty-one bioinclusions representing one basal Hexapoda: Collembola, five insect orders and a portion of a spider web were recovered from the Genoveva amber (Supplementary Data 3). Most specimens are complete and well preserved (Fig. 4). The assemblage includes 11 Diptera: Ceratopogonidae (3), Chironomidae (6), Scatopsidae (1) and Dolichopodidae (1); one Coleoptera: Tetratomidae; four Hymenoptera: Scelionidae (1), †Stigmaphronidae (1), and Hymenoptera indet. (2); one Trichoptera. All these previous examples are holometabolous insects. The remaining two belong to the Hemiptera: Aleyrodidae (1), and Hemiptera indet. (1), which are representative of paraneopteran insects.
Diptera are the most common insects found in the Genoveva amber, consistent with their prevalence in other Cretaceous amber deposits22. One male specimen of the family Ceratopogonidae could be identified; it belongs to the genus Archiaustroconops (Fig. 4a). Although the preserved specimens are male, Lower Cretaceous ceratopogonids are known to include haematophagous females that fed on vertebrates23. Six additional specimens belong to the family Chironomidae (Fig. 4b and Supplementary Fig. 4a) with at least two distinct morphotypes recognised. Their presence suggests the occurrence of freshwater bodies or persistently moist conditions within the palaeoforest. One further specimen represents a new species of Microphorites, Dolichopodidae (Fig. 4c and Supplementary Fig. 4a), closely related to M. extinctus from the Barremian Lebanese amber24.
The general habitus and preserved morphological characters of the Coleoptera specimen suggest that it belongs to the Eustrophinae of the Tetratomidae (Fig. 4f–g). The fossil record of Tetratomidae is notably sparse, with occurrences known almost exclusively from amber inclusions25. Additional details on the dipteran and coleopteran inclusions are provided in the Supplementary Data 3.
The families †Stigmaphronidae (Fig. 4h) and Scelionidae (Fig. 4i) were identified within the Hymenoptera; additionally, an undetermined hymenopteran specimen was found. The stigmaphronid specimen is incomplete, lacking the wings and portions of the body, but is easily identified by its flattened and flared apically tibiae and their strong distal spurs. The scelionid is complete and well preserved, showing the characteristic habitus and stigmal vein in the fore wing of this family. These small wasps likely acted as parasitoids of other insects, a behaviour documented in extant representatives26 and inferred for †Stigmaphronidae27. While scelionids are common in Cretaceous ambers28, stigmaphronids are relatively rare27.
The order Trichoptera (Fig. 4e) is only represented by one incomplete adult specimen. The wings are partially visible in the preparation and largely overlap each other, and the body features are obscure or not preserved. The typical dense setae of trichopterans on the wing membrane not modified into scales are well visible, but the reconstructed wing venation is not enough to make a taxonomic identification to the family level. The presence of this adult specimen indicates the existence of nearby water bodies, as larvae of this order are aquatic.
In addition to the previously described holometabolous insects, the Genoveva amber includes a single specimen of Aleyrodidae (Hemiptera: Sternorrhyncha) and another one considered as Hemiptera indet., the only representatives of the paraneopteran insects recovered so far. The aleyrodid is an adult preserved in a fragment of cloudy amber that partially hides its wings (Supplementary Fig. 4b). Like other sternorrhynchans, aleyrodids are phytophages. They are well represented in both Cretaceous and Cenozoic amber deposits and are known for their high diversity.
A portion of the spider web found is constituted by seven right-strand portions, ca. 5–7 μm thick, in an assemblage that suggests an orbicular web, because the strands are not in a random distribution and are in the same plane. Also, because two parallel strands, ca. 5 mm length each, are in contact with a strand crossing them, also ca. 5 mm length, in a relative perpendicular orientation in respect to both (Supplementary Fig. 4c). No prey is associated with this specimen, and the silk strand lacks the sticky droplets characteristic of orbicular spider webs, unlike some reported from Albian Spanish amber29.
Pseudoinclusions and bio-pseudoinclusions in Genoveva amber
Genoveva amber is rich in two types of pseudoinclusions, including mineralisations of iron sulphide (Supplementary Fig. 8a), and bio-pseudoinclusions in the manner of resin-in-sap-in-resin double emulsions or fossilised phloem sap (Supplementary Fig. 8d, e). These double emulsions containing phloem sap were first described from Albian amber of El Soplao (Spain)30, and have been observed in diverse Cretaceous ambers in the Northern Hemisphere.
Other bio-pseudoinclusions are mycelia of resinicolous fungi forming cortices in the amber lumps (Supplementary Fig. 8a)31,32. Although this type of fossil mycelia is very abundant in some Cretaceous ambers, in the Genoveva amber are rare. In cases where the hyphae penetrated the resin, they become relatively well preserved, and generally only the empty hyphal core is visible under optic microscopy (Supplementary Fig. 8b). However, in the Genoveva amber, some parts of the hyphal cores are filled with iron sulphide mineralisations (Supplementary Fig. 8c).
Genoveva botanical assemblage and amber age
No plant remains have been found as bioinclusions in the Genoveva amber. Nevertheless, a remarkable assemblage of pollen, spores and plant macrofossils are recognised in the Hollín Formation (Fig. 5). Ten dark fine-grained rock samples for pollen were collected along the 58 m thick stratigraphic log of Genoveva quarry (Supplementary Fig. 3). The macrofossil plant assemblage was collected mainly from the Genoveva quarry (Fig. 5a–m and Supplementary Fig. 3) and includes diverse fern allies and ferns (i.e., Selaginella, Acrostichum, Weichselia and Cladophlebis fronds) (Fig. 5a), cycadophytes (leaves of at least two taxa), araucariaceans, †cheirolepidiaceans, and cupressaceans. The Hollín Formation at the Genoveva quarry is particularly remarkable for preserving the earliest documented association of angiosperm leaves from north-western South America. All fossil leaves analysed under fluorescence microscopy also revealed a variety of epiphyllous fungi.
Fig. 5: Plant diversity (macrofossils and pollen grains) from the Hollín Formation at the Genoveva quarry.
a Selaginella. b–d Diverse fern pinnules. e Strap-shaped Araucariaceae leaf. f Cupressaceae branch. g Isolated cuticle of a probable Podocarpaceae leaf. h, i Isolated cuticle of Ginkgo-like leaf, note stomata with papillae. j Detail of †Cheirolepidiaceae leaf, note stomatal bands. k–m Diverse angiosperm leaves showing different types of venation and leaf shapes. n, o Araucariaceae pollen grains: Araucariacites australis, Balmeiopsis limbatus, respectively. p Angiosperm pollen: Retitricolpites operculatus. q Pollen grain with unknown affinity: Reyrea polymorphus. Photographed under epifluorescence in (h–j). Scale bars: 1 cm (c–f, l), 5 mm (a, b, k, m), 500 µm (g, h, j), 200 µm (i), 10 µm (n–q).
The taxonomic composition of the palynoflora (68 taxa in total) in the Genoveva quarry (Fig. 5n–q) is dominated by pteridophytes (42% of the taxa), followed by angiosperms (37%), gnetophytes (13%) and conifers (8%) (Supplementary Table 1, pollen taxonomic composition), while the palynoflora abundance is also dominated by pteridophytes (49%) followed by angiosperms (24%), conifers (21%) and gnetophytes (5%). Most of the conifer pollen abundance is dominated by araucariaceans (72%, Araucariacites australis and Balmeiopsis limbatus). The age of the Hollín Formation spans from the early Aptian to the middle Albian6,8,33,34,35. The age of the Hollín Formation at the Genoveva quarry, which corresponds to the Hollín Principal Member, is inferred based on the co-occurrence of Retitricolpites operculatus and Reyrea polymorphus, characteristic of palynological Zone 636, which is dated to the Albian.
Palaeoenvironment and palaeoclimate
The new palaeobotanical evidence suggests the presence of a diverse and humid, low-latitude forest in north-western Gondwana during the early Albian. Pollen and plant macrofossils show abundant ferns and fern-allies that likely grew in the understory and/or near water bodies, in a forest dominated by araucariacean resinous trees (Fig. 5). The overall palynological and plant macrofossil association found in the Genoveva quarry (Supplementary Table 1), particularly the high diversity of pteridophytes and the presence of moderately thick coal seams in the stratigraphic sequence (Supplementary Fig. 3), indicates a humid environment, similar to previous reports in other but less studied north-western tropical South American sites37,38,39.
The presence of relatively abundant chironomid flies and one trichopteran as bioinclusions—both insect groups with aquatic larval stages—further supports the interpretation of predominantly humid conditions during resin production and deposition. These results contrast with those from eastern tropical South America sites, where the extensively studied Crato Formation indicates arid climate conditions40,41,42,43,44. Moreover, several global climate models have inferred a precipitation gradient for the middle Cretaceous within the tropics of South America, with wetter conditions in the west and drier conditions in the east45,46.
No evidence of palaeofires has been found in the Genoveva quarry strata, supporting the palaeobotanical interpretation that the resin-producing forests were humid, with conditions unfavourable to fire. This is in contrast to what has been observed in the Northern Hemisphere3, where the presence of charcoal associated with Cretaceous amber is quite frequent.
Unlike many Cretaceous ambers from the Northern Hemisphere31, the Genoveva amber shows a scarce presence of resinicolous fungal mycelia (cortices), indicating that the equatorial resinous trees likely grew in soils with different humidity level. It is possible that the soils in this region were saturated with water, inhibiting the fungal growth, as has been inferred for Albian Ariño amber in Spain21. Despite the rarity of these Mesozoic resin-recycling microorganisms in the Genoveva amber, their identification marks the first record for South America.
Conclusions
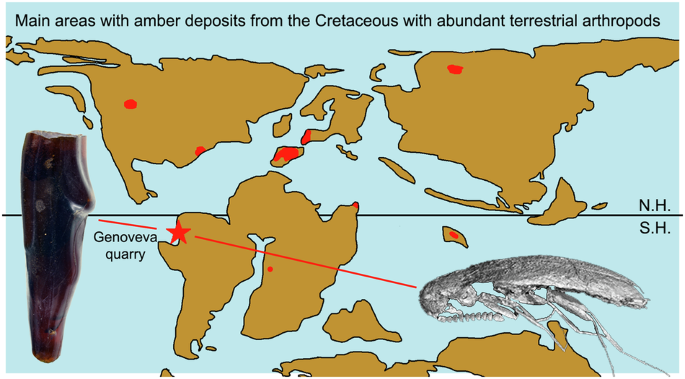
Cretaceous amber-containing insects and spider web remains is reported here from South America, a region where such inclusions had not been previously documented. This discovery and the associated plant remains in the amber-bearing rocks, enhance our understanding of the Gondwanan arthropod fauna and flora inhabiting forests along its western margin during a time interval of major ecosystem transformation. The Genoveva amber represents the largest Mesozoic amber deposit in South America and is one of the few deposits in the Southern Hemisphere with abundant amber during the Cretaceous Resinous Interval, making it critical for global studies of this Cretaceous phenomenon.
Future fieldwork exploration in the Hollín Formation of the Napo region is expected to yield new fossil bioinclusions which may improve our understanding of the biogeographic relationships between the South American fauna and flora and those of other Gondwana regions, such as Antarctica, South Africa, Australia, and New Zealand, where amber is also present and need to be investigated.