
Fe₃O₄ and PGRs elevate plant growth
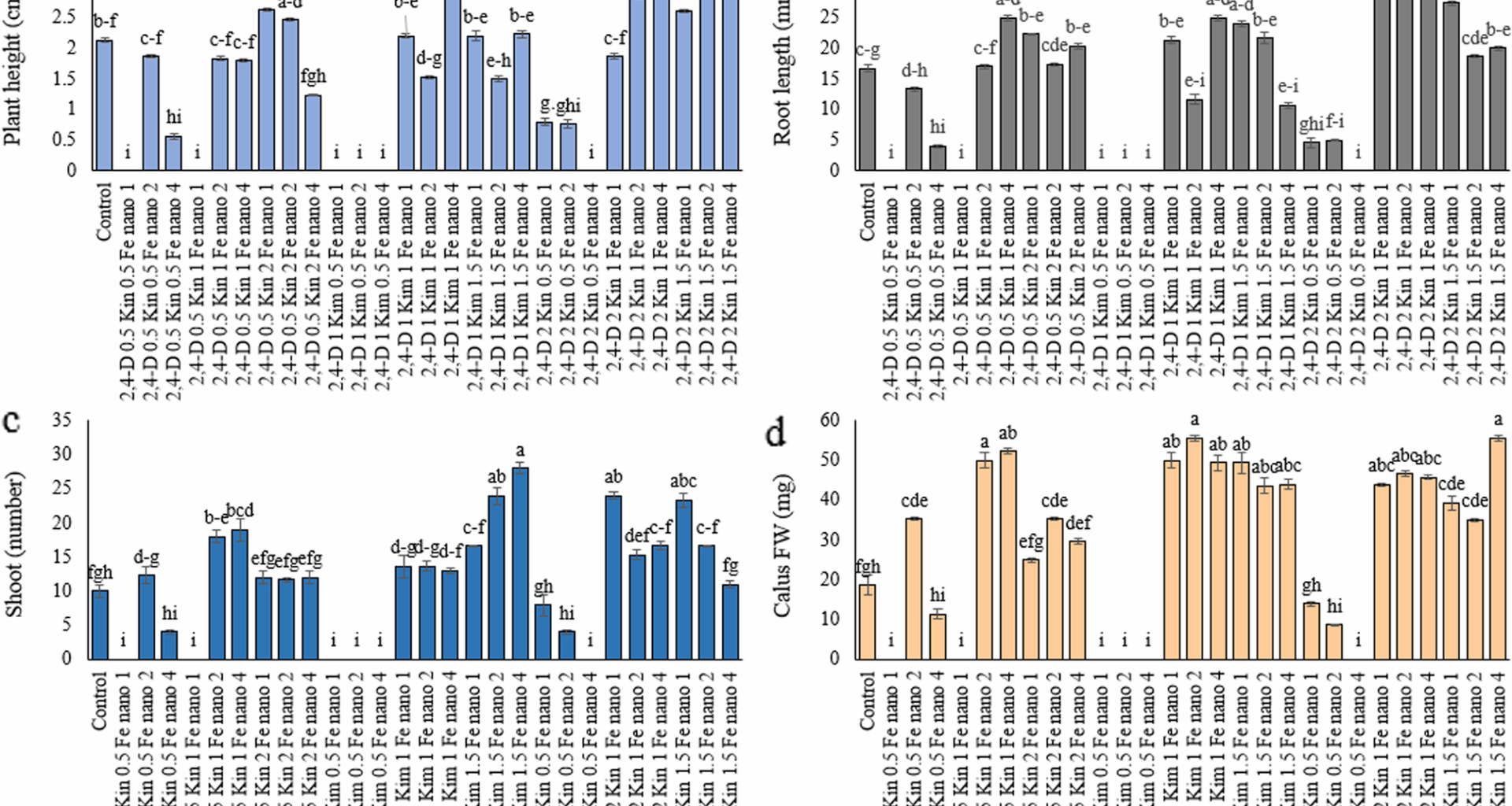
Several treatments including 2,4-D (0.5 to 2 mg/L), kinetin (0.5 to 1 mg/L), and the Fe₃O₄ nanoparticles (1 to 4 mg/L) produced reductions in plant height when compared to the control treatment. The treatments that had the greatest reductions in height were 2,4-D 0.5 mg/L + kinetin 0.5 mg/L + Fe₃O₄-NP 1 mg/L, 2,4-D 0.5 mg/L + kinetin 1 mg/L + Fe₃O₄-NP 1 mg/L, 2,4-D 1 mg/L + kinetin 0.5 mg/L + Fe₃O₄-NP (1, 2, and 4 mg/L), and 2,4-D 2 mg/L + kinetin 0.5 mg/L + Fe₃O₄-NP 4 mg/L (Fig. 1a). The decrease in plant growth seen in certain treatments when compared to the control is in line with other research showing that hormone abnormalities or improper concentrations of nanoparticles can have a negative impact on plant development [15, 16]. The combination of 2 mg/L 2,4-D, 1 mg/L kinetin, and 4 mg/L Fe₃O₄-NP increased plant height by 59.6% compared to the control, representing the highest increase observed (Fig. 1a). Iron nanoparticles serve as a source of micronutrients and increase plant metabolism by increasing the activity of enzymes involved in cellular respiration and photosynthesis [17, 18]. In addition, growth hormones including kin and 2,4-D can promote root and shoot development by affecting cell division and differentiation [6, 19]. Ali and Alwani Bahm [20] analyzed the impact of a combination of 4 mg/L 2,4-D and 4 mg/L kin on the Trigonella foenum-graecum L. and noted a statistically significant increase in plant height along with biomass accumulation. In another investigation, Muthu et al. [21] applied 2,4-D at 5 µM mg/L and kin at 10 µM on Glycine max (soybean) seedlings and documented better root and shoot growth than controls. Likewise, Ullah et al. [22] treated Oryza sativa L., with 15 ppm FeO-NPs alone, and observed increases in plant height.
Fig. 1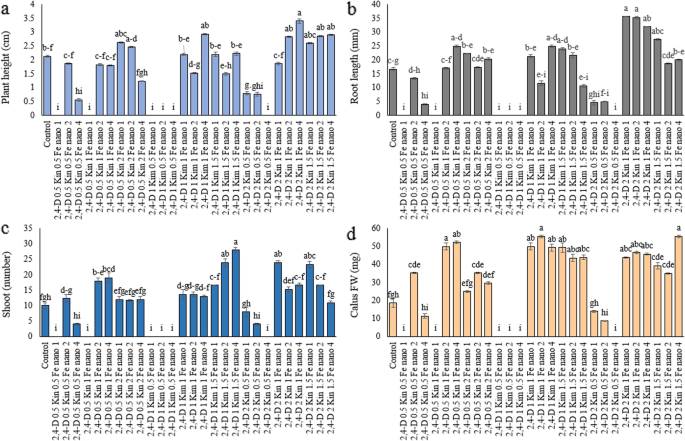
Effect of treatment compounds, Kin, 2,4-D and Fe₃O₄-NP on Hypericum perforatum L.: (a) plant height, (b) root length, (c) shoot number, and (d) callus FW. Data represent mean ± SE (n = 3). Different letters indicate significant differences between treatments based on LSD test (P ≤ 0.05)
Root length and shoot number boost with PGRs and Fe₃O₄-NP
The significant increase in root length in the combined treatment of 2 mg/L 2,4-D, 1 mg/L kin, and Fe₃O₄-NP at concentrations of 1 and 2 mg/L (114.04 and 112.06% compared to the control, respectively) indicates the synergistic effect of these compounds on root growth (Fig. 1b). The combined treatment of PGRs and Fe₃O₄-NP had a positive effect on increasing the number of shoots, although this effect was not observed in all treatments. It is noteworthy that the treatment containing 1 mg/L 2,4-D, 1 mg/L kin and 4 mg/L Fe₃O₄-NP caused a 180% increase in the number of shoots compared to the control (Fig. 1c). Previous studies have shown that using growth regulators like 2,4-D and kin together can positively influence root differentiation and growth [23,24,25]. Additionally, Fe₃O₄-NP boosts nitrogen metabolism and enhances photosynthetic efficiency thanks to their large specific surface area and their ability to slowly release iron ions [26]. Using iron in its nano form is a much an effective way to boost root growth since plant cells absorb it better [27]. Iron plays a crucial role in the makeup of enzymes that are vital for plant growth, and without it, root development can really suffer [28]. The present investigation matches the findings of Tombuloglu et al. [18], since the study showed that iron nanoparticles combined with cytokinins produced enhanced root system growth and more shoot emergence in tomato seedlings. The research by Yousaf et al. [29], demonstrated that Fe₃O₄-NP together with growth regulators promoted higher biomass growth and improved root development in corn plants. In Oryza sativa L., nanogreen iron oxide (FeO-NPs) with sizes of 2–7.5 nm effectively enhanced tissue culture performance. Supplementation of MS media with 15 ppm FeO-NPs significantly reduced callus contamination by 80%. FeCl₃-based FeO-NPs promoted rapid callus induction (72%) and regeneration (43%), whereas FeSO₄-based FeO-NPs led to greater callus biomass (516%), diameter (300%), shoot number (200%), and root formation (114%) [22].
Boost in callus weight by PGRs and fe₃o₄ nano
The combined application of plant growth regulators (PGRs) and Fe₃O₄-NP increased the callus FW compared to the control treatment (Fig. 1 d). Specifically, plants using the combination of 1 mg/L 2,4-D, 1 mg/L kin and 2 mg/L Fe₃O₄-NP and treatment of 2 mg/L 2,4-D, 1.5 mg/L kin and 4 mg/L Fe₃O₄-NP resulted in 198.28% callus FW gain compared to control. 2,4-D is a synthetic auxin that enhances cell division [30], while kin, is a cytokinin that promotes cell division in meristematic zones and delays cell senescence [31]. Fe₃O₄-NP presence can enhance enzyme activities (e.g., POD, CAT) by improving cellular iron uptake and related metabolic processes [32]. Iron nanoparticles stimulate callus development through their small size and high surface-to-volume ratio by improving cell wall permeability [33]. Research evidence from different experiments demonstrates the same conclusion; the combination of iron nanoparticles with plant growth regulators effectively promoted callus expansion in Catharanthus roseus according to Sepehri et al. [34], Iron nanoparticles served to improve wheat tissue growth through their effects on photosynthesis and chlorophyll production according to Feng et al. [26]. The study by Zahmatkesh et al. [35], demonstrated that combining 2,4-D (1 mg/L) with kin (0.5 mg/L) produced significant increases in H. perforatum callus fresh weight and dry weight. The experiment by Shasmita et al. [6] demonstrated that 2,4-D alone produced successful callus formation in H. perforatum while adding kin to the mixture produced better callus fresh weight results and enhanced hypericin production. The application of low-concentration Fe₃O₄-NP combined with PGRs in H. perforatum resulted in significant improvements in callus weight together with colorimetry measurements and enzymatic activity according to Kralova and Jampilek [36]. They attributed this phenomenon to stimulation of cellular metabolism via increased iron availability.
Occurrence of black spots on leaves
Most treatments of either PGRs and Fe₃O₄-NPs did not induce the formation of black nodules on leaves from regenerated shoots and there were no nodules in any of the treatments with 2,4-D and the concentrations of 0.5 or 1 mg/L, kinetin and the concentrations of 0.5 or 1 mg/L, and in the applications of Fe₃O₄-NPs at 1, 2 or 4 mg/L. There was a slight (but not statistically significant) increase in nodule development for a few combinations at higher concentrations of kinetin and/or Fe₃O₄-NPs, e.g., 2,4-D (0.5–2 mg/L) with kinetin (1–1.5 mg/L) and Fe₃O₄-NPs (2–4 mg/L). These results are illustrated in Fig. 2. Black nodules are either hypericin rich glands that are both natural and desirable. In our study, the black nodules were primarily associated with tissue browning a suggesting to be favourable hypericin accumulation. Treatments with 2,4-D, kinetin, and Fe₃O₄ nanoparticles resulted in greater callus growth, less tissue darkening and increased chlorophyll content, which suggests improved metabolic function and potential reduction in oxidative damage [37]. The investigation assessed the influence of kin with 2,4-D and Fe₃O₄-NP on H. perforatum L. callus growth and development (Fig. 3a–g). Prior research has shown that Fe₃O₄ nanoparticles have antifungal and antibacterial activity [38, 39] and in combination with plant growth regulators, can stimulate plant defense mechanisms and mitigate cellular stress indicators [18]. Although the biosynthesis of secondary metabolites is genetically regulated, their actual production is strongly influenced by environmental conditions. Metabolic and morphogenetic changes can alter the levels of secondary metabolites by modulating the expression of specific plant genes in response to environmental factors. In addition, elicitors may trigger the activation of new genes that stimulate enzymes and, consequently, various biosynthetic pathways, leading to the formation of secondary metabolites. For example, polyketide synthase (PKS) plays a central role in the biosynthesis of hypericin and hyperforin. Similarly, phenylalanine ammonia lyase (PAL) is a key enzyme whose upregulation increases the accumulation of phenolic compounds. Under mild stress conditions, these precursors can be converted into anti-inflammatory metabolites such as hypericin, thereby enhancing the plant’s resistance to stress.Pretto (2006) reported that in Hypericum perforatum cell suspension cultures, treatment with 250 µM jasmonic acid induced higher levels of hypericin compared to lower concentrations (50 or 100 µM). Moreover, another study demonstrated that reduced shading, which increases sunlight exposure and consequently daytime heat in natural habitats of H. perforatum, also contributed to higher hypericin production.Based on these findings, the present study examined Fe nanoparticles as potential elicitors in tissue culture. The results indicated that these nanoparticles could activate genes involved in hesperidin biosynthesis by enzymatic stimulation, leading to an increased accumulation of this compound (Fig. 2). Therefore, compared with plants grown under natural conditions, it can be suggested that when mild stresses are combined with the activation of biosynthetic genes for secondary metabolites, the production of such compounds may also be enhanced in vivo.
Fig. 2
Effect of treatment compounds, Kin, 2,4-D and Fe₃O₄-NP on Hypericum perforatum L.: NO. of black/leaves. Data represent mean ± SE (n = 3). Different letters indicate significant differences between treatments based on LSD test (P ≤ 0.05)
Fig. 3
Effect of treatment compounds, Kin, 2,4-D and Fe₃O₄-NP on Hypericum perforatum L.: (a) callus induction, (b) callus growth, (c) callus formation, (d) shoot induction (e) shoot frowth (f) seedling formation (g) intact plant, (f) plant acclimatization. Data represent mean ± SE (n = 3). Different letters indicate significant differences between treatments based on LSD test (P ≤ 0.05)
Kin, 2,4-D & Fe3O4 nano: callus and rooting percentage
In this research study the percentage of callus and root formation, from the combined treatments of kinetin(Kin), 2,4-D, and Fe₃O₄ nanoparticles (Fe₃O₄-NP), was significantly increased from the control (Fig. 4a, b). Specifically, the treatment of 2,4-D at 0.5 mg/L, kinetin at 1 mg/L, and Fe₃O₄-NP at 4 mg/L resulted in a 62.26% increase in callus formation, while the treatment of 2,4-D at 1 mg/L, kinetin at 1.5 mg/L, and Fe₃O₄-NP at 1 mg/L resulted in a 71.66% increase in root formation, when compared to the control (Fig. 4a, b). The effects of these compounds on medicinal plants have been demonstrated through various studies. Also, the research performed by Rastegaran et al. [40] demonstrated that Fe₃O₄-NP enhances callus development alongside root formation in chamomile (Matricaria chamomilla) through better iron supply and improved metabolic enzyme performance. Another study by Amiri et al. [41] demonstrated how borage (Echium amoenum) growth hormones combined with iron nanoparticles enhanced root production while also improving callus quality. Certainly, phytohormones like auxins and cytokinins are critical for in vitro caulogenesis and morphogenesis, as reported by several studies that revealed detrimental effects of excess or prolonged application on plant health [42,43,44]. In Study Arain et al. [45], excessive treatment with 2,4-D in Saccharum officinarum caused somaclonal changes, oxidative stress, and abnormal proliferative callus growth. Similarly, kinetin and principally other cytokinins used at high concentrations may alter endogenous hormone balances, lead to rooting difficulties [46]. These complications suggest that while exogenous hormones play an important role in morphogenesis, concentration and combination are critical in reducing stress and developmental variations [47, 48]. In a report on bell pepper, it was determined that the highest average proliferation for callus was recorded at a concentration of 6.0 ml/liter of MgNPs in MS culture medium, reaching 6.1 g/m2 [49].
Fig. 4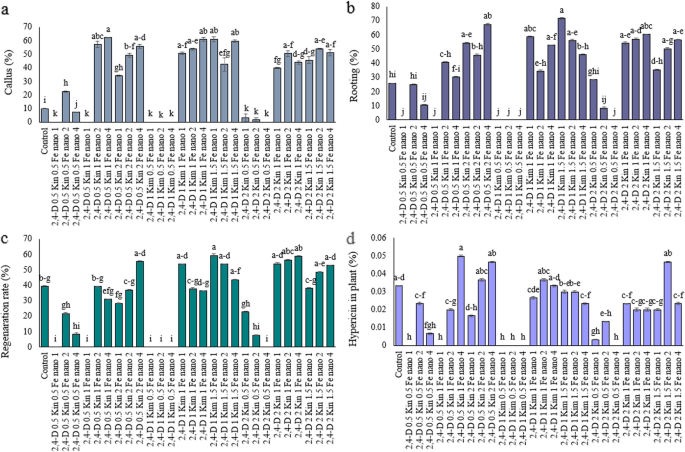
Effect of treatment compounds, Kin, 2,4-D and Fe₃O₄-NP on Hypericum perforatum L.: (a) callus percentage, (b) rooting percentage, (c) regeneration rate, and (d) hypericin in plant. Data represent mean ± SE (n = 3). Different letters indicate significant differences between treatments based on LSD test (P ≤ 0.05)
Regeneration rate: increase with Kin, 2,4-D & Fe3O4 Nano combination
The treatment combinations generally had a positive effect on the percentage of regeneration rate, but in some cases this effect was reduced. It is noteworthy that the treatment of 2,4-D 1 mg/L, kin 1.5 mg/L and different concentrations have independent significantly increased the regeneration rate by 50.85% compared to the control (Fig. 4c). These findings highlight that specific nano-hormonal combinations can act synergistically to enhance growth and metabolite production, an approach that can benefit medicinal plant tissue culture. Research by Palmer and Keller [50] showed that 2,4-D treatment independently triggered callus formation in H. perforatum but kin addition significantly enhanced both callus development and regeneration. The improved hormonal equilibrium together with cell division stimulation generated this effect. The research Bal et al. [51] discovered that Fe₃O₄-NP treatment independently strengthened antioxidant enzyme function to improve root growth in H. perforatum. Culture medium containing 0.5 mg/L, 1 mg/L, and 1.5 mg/L of Fe3O4 NPs increased the regeneration rate of flax plants [52]. Using the three applications of Kin, 2,4-D, and Fe₃O₄-NPs resulted in a high rate of callus and root formation, but in vitro regeneration using these hormones has the potential for adverse events that need to be considered. For example, using 2,4-D or kinetin at either prolonged or high packet volumes may result in somaclonal variation, oxidative stress, abnormal morphology of callus, or tissue vitrification. Therefore, a high regeneration rate isn’t necessarily a predictor of genetic fidelity. Any regenerated plants should be evaluated for homogeneity and similarity to an in vivo plant using molecular markers (e.g., RAPD, ISSR, AFLP) or cytogenetic analyses [53,54,55].
Powerful boost of hypericin via 2,4-D, Kinetin & Fe3O4 nano
The improvement in the amount of hypericin in the plant was particularly significant with the treatment of 2,4-D 0.5 mg/L, kin 1 mg/L and Fe₃O₄-NP 4 mg/L, 66.66% increase compared to the control plants (Fig. 4d). Also, the treatments of 2,4-D 0.5 mg/L, kin 2 mg/L and Fe₃O₄-NP 4 mg/L and 2,4-D 2 mg/L, kin 1.5 mg/L and Fe₃O₄-NP 2 mg/L showed a 39.39% increase in hypericin compared to the control, respectively, and these two treatments did not differ significantly from each other (Fig. 4d). The active component hypericin forms part of the healing process in H. perforatum while growing production improves the economic value and medical benefits of the product [1]. In the work of Al-Khayri et al. [56] H. perforatum callus cultures synthesized more hypericin in the presence of 2,4-D and kin applied simultaneously and the maximum hypericin content was reached at a lower concentration of 2,4-D (0.5–1 mg/L) combined with kin (1–2 mg/L). Aminizadeh et al. [57] showed that Fe₃O₄-NP applied to Lepidium sativum led to increased activity of antioxidant enzymes and higher production of phenolic compounds. The mode of the effects of H. perforatum on the enhancement of hypericin content is comparable to Fe₃O₄-NP for their increases of activity of antioxidant enzymes and production of phenolic compounds shown in Lepidium sativum plant [57]. The researchers Abed et al. [58] reported that application of iron nanoparticles along with 2,4-D and BAP on Catharanthus roseus plants improved production of alkaloids. However, the results are also indicative of the fact that not only nanoparticles interact with PGRs and thereby enhance the availability of the minerals, but they also enhance the tissue redox status ultimately enhancing the alkaloids biosynthesis.
Treatment-Induced changes in phenol and flavonoid levels
The increase in phenolic content by the treatments 2,4-D 0.5 mg/L + Kin 1 mg/L + Fe₃O₄-NP 4 mg/L, 2,4-D 0.5 mg/L + Kin 2 mg/L + Fe₃O₄-NP 1 mg/L, 2,4-D 1 mg/L + Kin 1 mg/L + Fe₃O₄-NP 4 mg/L, 2,4-D 1 mg/L + Kin 1.5 mg/L + Fe₃O₄-NP 1 mg/L, 2,4-D 2 mg/L + Kin 1 mg/L + Fe₃O₄-NP 2 mg/L, 2,4-D 2 mg/L + Kin 1 mg/L + Fe₃O₄-NP 4 mg/L, and 2,4-D 2 mg/L + Kin 1.5 mg/L + Fe₃O₄-NP 1 mg/L was 72.21%, 75.38%, 80.14%, 64.28%, 64.25%, 64.27%, and 75%, respectively, compared to the control (Fig. 5a). Chakraborty et al. [59] found that the use of 2,4-D in Withania somnifera during in vitro cultivation improved the synthesis of phenolic compounds and other secondary metabolites as well as quality of callus. Also, Emamverdian et al. [60] found that applying Fe₃O₄-NP along with growth regulators like kin increased the phenol content and activity of POD and CAT enzymes that is related to enhancement of the plant defense status in stress conditions. The combined application of 2,4-D and kin increased the contents of total phenols, flavonoids and the activity of antioxidant enzymes in the in vitro cultures of Ocimum basilicum L [61]. Triggered phenol production was also observed in in vitro culture medium of Artemisia annua L. upon Fe₃O₄-NP and kin treatment [62]. Synergistic application of kin and Fe₃O₄-NP enhanced the content of phenolic and flavonoid metabolites in emon balm (Melissa officinalis L.). Moreover, the co-administration of Fe₃O₄-NP and kin in Thymus vulgaris L. enhanced the total phenol production and reduced oxidative stress [63]. In a study, the synergy of a nitrophenol-based biostimulant nanocomplex caused an increase in phenols and flavonoids in tomatoes [64].
Fig. 5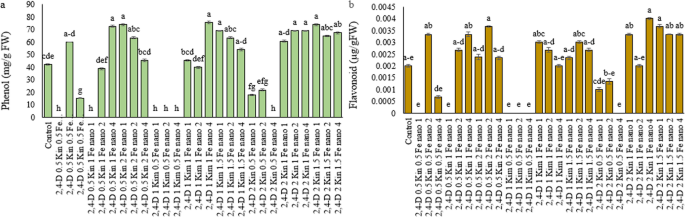
Effect of treatment compounds, Kin, 2,4-D and Fe₃O₄-NP on Hypericum perforatum L.: (a) phenol, and (b) flavonoid. Data represent mean ± SE (n = 3). Different letters indicate significant differences between treatments based on LSD test (P ≤ 0.05)
Evaluation of secondary metabolites
The synthesis of important secondary metabolites in Hypericum perforatum was significantly enhanced by the joint addition of 2,4-D, kin, and Fe₃O₄-NP. The most abundant compounds were hypericin and hyperforin (24.6 ± 0.027 and 18.2 ± 0.001 (Table 1), respectively indicating more effective therapeutic usefulness. Identified compounds accounted for (93.36%) was also affected by other bioactive molecules including ß-patchoulene, hexadecanol and naphthalene. High data precision (r = 0.96; CV = 4.22%) confirmed the reliability of these discoveries. This work with potential use in pharmaceutical and agricultural sectors indicates the synergistic effects of PGRs with Fe₃O₄-NP to enhance the phytochemical contents in H. perforatum (Table 1). Figure 6 represents the GC graph of treatment compounds 2,4- D 1 mg/L, Kin 1 mg/L, Fe₃O₄-NP 4 mg/L. In a study of St. John’s wort (H. perforatum L.), the compounds hypericin, pseudohypericin, hyperoside, rutin, quercitrin, and hyperforin were obtained [65].
Table 1 Evaluation of secondary compounds in the best treatment combination including 2,4-D 1 mg/l, Kinetin 1 mg/l, and Fe₃O₄-NP 4 mg/lFig. 6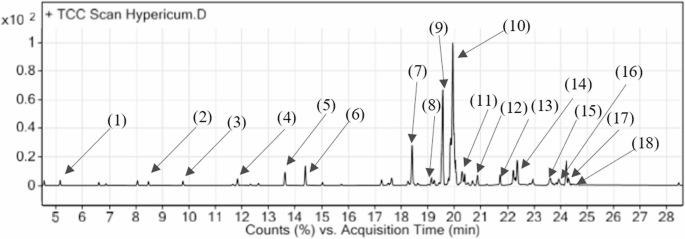
GC Chromatograph of the treatment compounds 2,4-D 1 mg/L, kinetin 1 mg/L, and Fe₃O₄-NP 4 mg/L
Multivariate trait analysis (HCA and PCA)
The results show statistically significant positive correlations in treatments with 2,4-D, kin, Fe₃O₄-NP and all growth parameters and secondary metabolites measured. The treatment of 2,4-D 0.5 mg/L + Kin 2 mg/L + Fe₃O₄-NP 4 mg/L showed significantly higher hypericin content and rooting percentage and regeneration (Fig. 7a), which indicated the strong stimulatory effect 2,4-D, kin and Fe₃O₄-NP on growth and metabolite production. Likewise, 2,4-D 1 mg/L + Kin 1.5 mg/L + Fe₃O₄-NP 1 mg/L improved regeneration and rooting rates, thereby providing support for its role in organ development and cell regeneration (Fig. 7a). The number of shoots was significantly higher with 2,4-D 1 mg/L + Kin 1.5 mg/L + Fe₃O₄-NP at 2 and 4 mg/L, indicating alterations in branching and structural growth. Additionally, hypericin production showed a positive relationship with 2,4-D 2 mg/L + Kin 1.5 mg/L + Fe₃O₄-NP mg/L and 2,4-D 0.5 mg/L + Kin 1 + Fe₃O₄-NP 4 mg/L supportive of their influence on metabolite pathways (Fig. 7a). Similarly, root length and plant height had positive relationships with particular treatments indicating improved uptake producing greater overall plant growth.
Fig. 7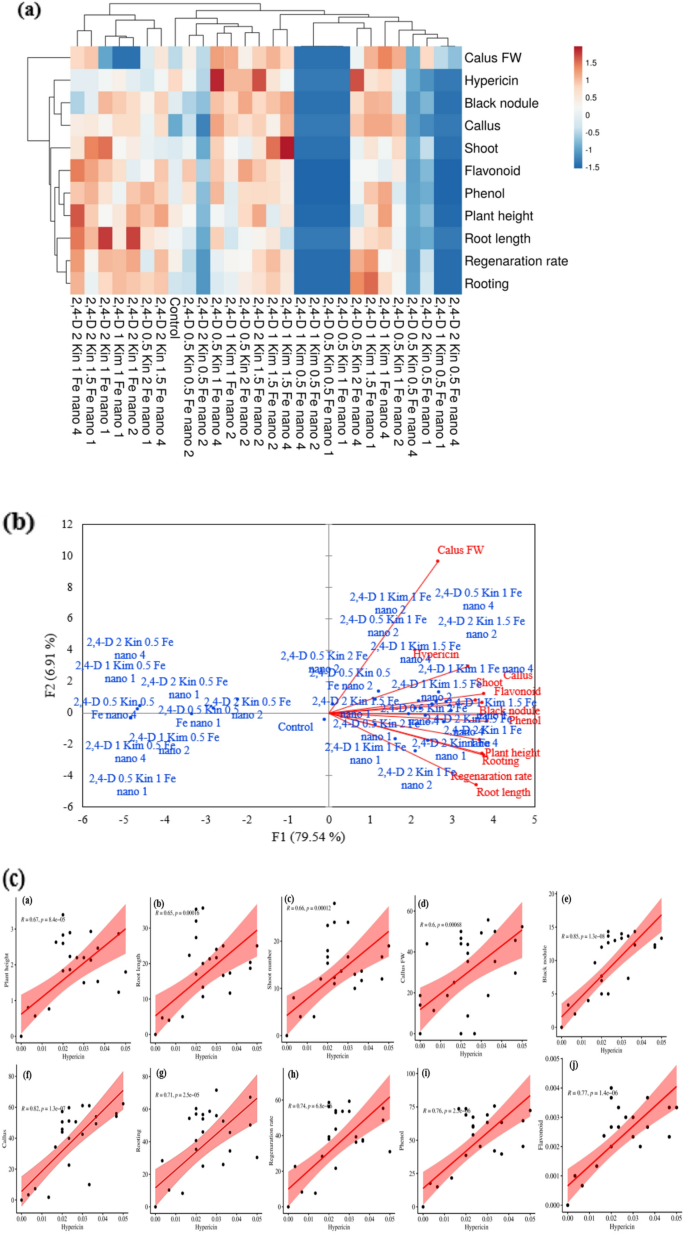
Effect of treatment compounds, Kin, 2,4-D and Fe₃O₄-NP on Hypericum perforatum L. (a) Hierarchical clustering analysis (HCA) of Pearson’s correlation coefficient (r) values of variable traits, where the color scale that indicates r coefficient values (r = 1.5 to −1.5) indicates positive (red) and negative (blue) correlations. (b) Principal component analysis (PCA). (c) Pearson correlation coefficient scatter plot. Pearson correlation coefficient was used to reflect the linear-related degrees of two variables. It’s range is −1 to 1.1 depicts totally positive correlated; −1 depicts totally negative correlated; 0 depicts no linear correlation
In this PCA analysis, treatments with 2,4-D (1, 0.5, and 2 mg/L), plus kin (0.5, 1, 1.5, and 2 mg/L), and Fe₃O₄-NP at different concentrations (1, 2, and 4 mg/L) had positive and significant correlations with different growth parameters and secondary metabolites (Fig. 7b). In particular, the treatment combinations of 2,4-D 1 mg/L with Kin 1 and 1.5 mg/L and Fe₃O₄-NP 2, 4 and 1 mg/L; 2,4-D 0.5 mg/L with Kin 1 and 2 mg/L and Fe₃O₄-NP 2 and 4 mg/L increased callus fresh weight, hypericin content, callus formation percentage, number of shoots and phenolic and flavonoid contents and decreased of black nodule accumulation. Conversely, the treatment combinations of 2,4-D 2 mg/L with Kin 1 mg/L and Fe₃O₄-NP 1, 2 and 4 mg/L and 2,4-D 1 mg/L with Kin 1 mg/L and 2,4-D 2 + Kin 1.5 + NP 1 mg/L had positive and significant correlations with root length, rooting percentage, rooting percentage and plant height (Fig. 7b). The findings highlight the different concentrations of treatment compounds have an independent effect on enhanced growth parameters and increased production of active metabolites and they combine as a synergistic formulation that can be beneficial for enhanced tissue culture and medicinal plant production.
Pearson correlation analysis
Pearson correlation analysis revealed a positive and significant association between hypericin contents and most of the morphological and biochemical traits. Most notably, the highest correlation coefficient observed was hypericin and black nodule formation (r = 0.85) and callus formation percentage (r = 0.82) which both signify the roles of hypericin in tissue differentiation and morphogenic processes (Fig. 7c). A significant correlation was also found for hypericin and rooting (r = 0.71), regeneration (r = 0.74), height (r = 0.67), root length (r = 0.65) and number of shoots (r = 0.66), which signifies simultaneous effects on plant growth. Biochemical data showed a strong correlation with phenolic compounds (r = 0.76) and flavonoids (r = 0.77), and may suggest a relationship between hypericin metabolism and the production of antioxidant compounds (Fig. 7c). This suggests that hypericin accumulation is positively linked with overall secondary metabolite production and antioxidant capacity in the cultures.