

The adenovirus penton base and HER3-Targeted HPK nano-capsids
Here, Cedars-Sinai Medical Center synthesize foundational adenovirus biology with key advances to present a unified model of penton base structure, function, and translational reprogramming
AUTHOR(S): Rebecca Benhaghnazar1, Thomas Mota1, Amirhesam Babajani1, Sun Woo Kim1, Giorgia Guerra1, Joseph Aceves1,2, Nelyda Gonzalez- Almeyda1, Kristin Ishaya1, Abby Wiesenthal1, Ravinder Abrol2, Lali Medina Kauwe1*
Department of Biomedical Sciences, Cedars-Sinai Medical Center, Los Angeles, CA, USA
Department of Chemistry and Biochemistry, California State University, Northridge, CA, USA
* Corresponding author
Overview and Viral Foundations
Summary
Efficient intracellular delivery – specifically escape from the endosomal compartment into the cytosol – remains one of the most formidable challenges in nucleic acid and macromolecular therapeutics.1 Viruses solve this problem through evolutionarily refined mechanisms that integrate receptor targeting, intracellular trafficking, and membrane disruption.2 Among these viral solutions, the adenovirus penton base stands out as a structurally discrete yet biologically powerful capsid protein that orchestrates internalization and endosomal escape.
Over the past two decades, mechanistic and translational studies have demonstrated that the penton base is not merely a structural support for adenovirus fibers, but a functionally autonomous, endosomolytic module. These discoveries enabled the rational engineering of HerPBK10 (HPK) – a HER3-targeted, penton base-derived delivery platform capable of self- assembling with nucleic acids into nano-capsids and mediating pH-triggered endosomal lysis.
Here we synthesize foundational adenovirus biology with key advances to present a unified model of penton base structure, function, and translational reprogramming.
1. Adenovirus penton base: Structural context
Adenoviruses are non-enveloped, double-stranded DNA viruses with an icosahedral capsid composed primarily of hexons and penton complexes at each of the twelve vertices. Each penton complex consists of a pentameric penton base anchoring a trimeric fiber protein, forming the primary interface between virus and host cell (Fig. 1).3-4
Each penton base monomer (~60 kDa) assembles into a homopentamer with a central pore and extended, flexible loops. High-resolution crystallography and cryo-electron microscopy revealed that one of these loops contains a conserved Arg-Gly-Asp (RGD) motif, enabling interaction with cellular αv integrins.5-7
The integrin binding region of the wild type penton base is naturally positioned in a sterically shielded location at the base of the fiber on the viral capsid surface, which helps the virus evade immune detection of these sites and prevents immune mediated interference with receptor interactions.32
2. Role in viral entry
Adenovirus entry is a multi-step process (Fig. 1).8 Initial attachment is mediated by fiber binding to primary receptors such as CAR, CD46, or desmoglein-2, depending on serotype.9-11
Subsequent engagement of αvβ3 and αvβ5 integrins by the penton base RGD motif activates intracellular signaling cascades that promote clathrin-mediated endocytosis.12-14
After internalization, virions traffic to early endosomes, where partial capsid disassembly occurs.15-17 Although internal protein VI contributes to membrane penetration, multiple studies demonstrated that the penton base itself possesses intrinsic membrane-destabilizing activity, capable of facilitating cytosolic access independently.18-24
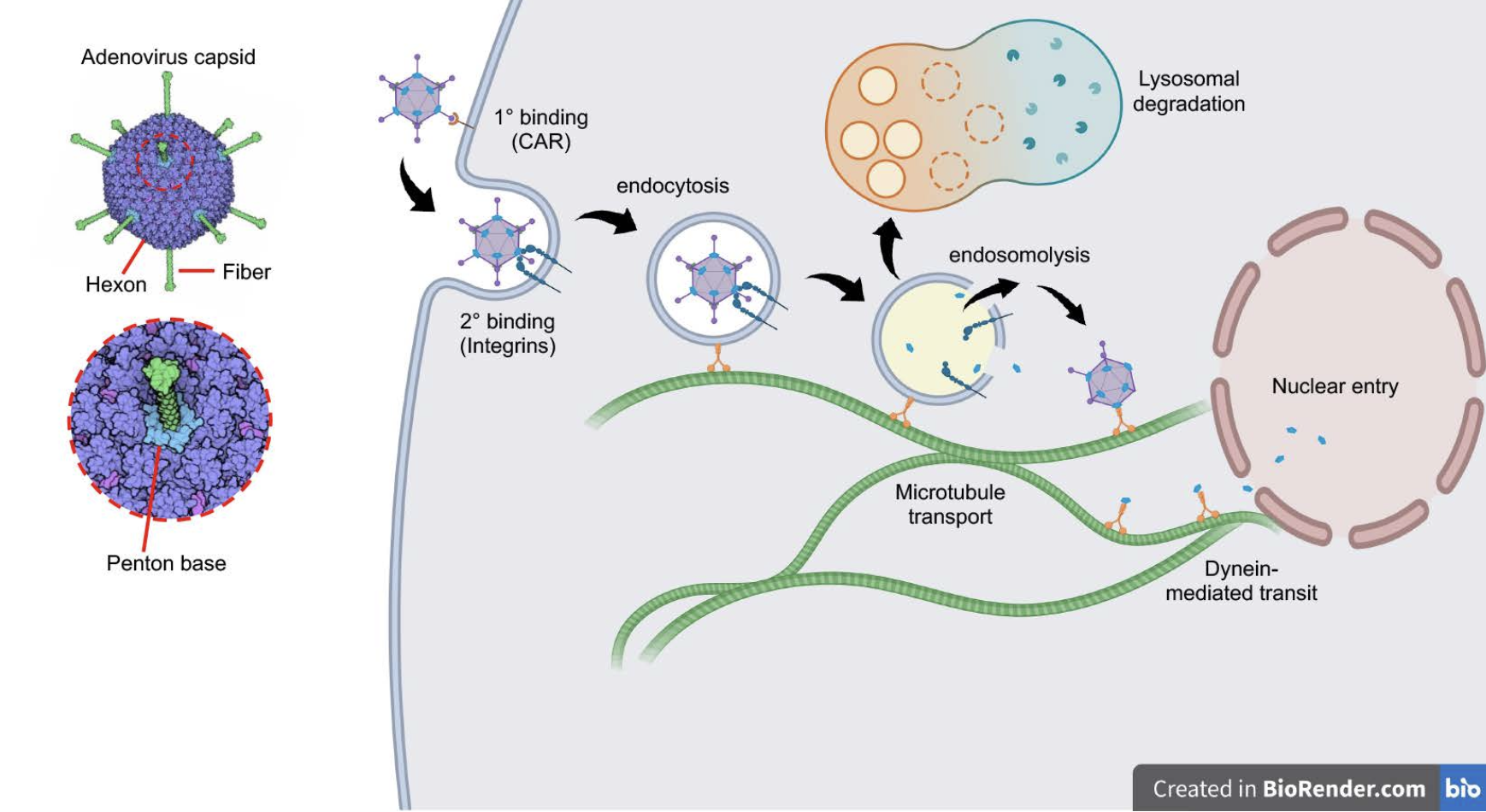 Fig. 1. Early stages of adenovirus infection. Schematic depicts the initial binding of adenovirus serotype 5 (Ad5) to its primary receptor, the coxsackievirus-adenovirus receptor (CAR) followed by secondary binding to alpha-v integrins, triggering integrin-mediated endocytosis. Endosome acidification triggers capsid-mediated disruption of the endosomal membrane, allowing escape of the virus into the cytoplasm and evasion of lysosomal degradation. Stepwise dismantling of the viral capsid during uptake enables soluble capsid components to hijack dynein-mediated transport systems, leveraging microtubule-supported trafficking toward the nuclear envelope and passage through nuclear pores. Adenovirus schematic is from Molecule of the Month, David Goodsell (2010) http://doi.org/10.2210/rcsb_pdb/mom_2010_12
Fig. 1. Early stages of adenovirus infection. Schematic depicts the initial binding of adenovirus serotype 5 (Ad5) to its primary receptor, the coxsackievirus-adenovirus receptor (CAR) followed by secondary binding to alpha-v integrins, triggering integrin-mediated endocytosis. Endosome acidification triggers capsid-mediated disruption of the endosomal membrane, allowing escape of the virus into the cytoplasm and evasion of lysosomal degradation. Stepwise dismantling of the viral capsid during uptake enables soluble capsid components to hijack dynein-mediated transport systems, leveraging microtubule-supported trafficking toward the nuclear envelope and passage through nuclear pores. Adenovirus schematic is from Molecule of the Month, David Goodsell (2010) http://doi.org/10.2210/rcsb_pdb/mom_2010_12
Translational re-engineering of the penton base
3. Engineering HerPBK10 / HPK
HerPBK10 (HPK) is a recombinant fusion protein designed to preserve the penton base’s membrane activity while introducing receptor specificity and cargo binding (Fig. 2).25-26 HPK comprises:
A heregulin-derived HER3-binding ligand,
A penton base–derived oligomerization and membrane-penetrating domain, and
A polylysine (K10) motif for electrostatic nucleic acid binding.
HPK internalization is independent of CAR and integrins25 and proceeds via HER3-mediated endocytosis,27-28 converting a broadly tropic viral mechanism into a tumor-selective delivery strategy.
4. HER3 targeting
HER3 (ErbB3) is overexpressed in many epithelial malignancies and plays a critical role in therapy resistance through heterodimerization with other ErbB receptors.29 HPK exploits this biology to achieve selective uptake in HER3-positive tumors, with minimal accumulation in normal tissues.27-28, 30
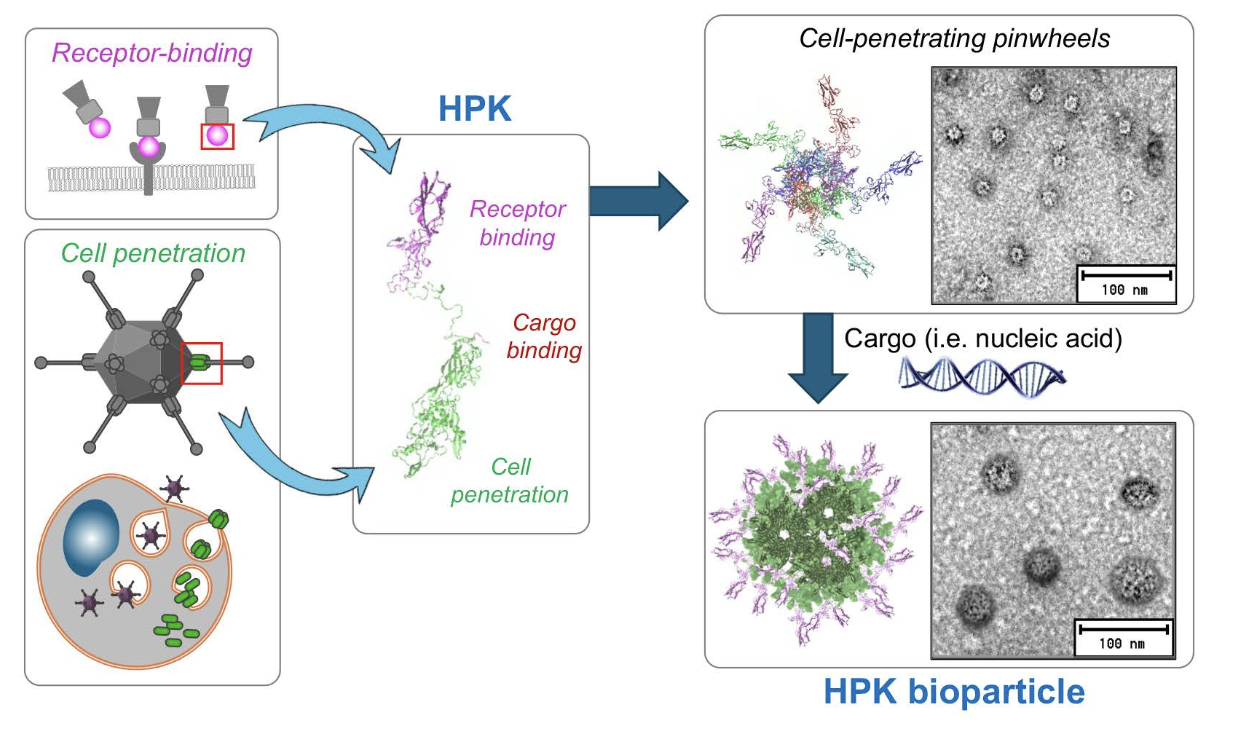
Fig. 2. HerPBK10 (HPK). Schematic depicts the combination of the receptor-binding region of heregulin alpha with the penton base sequence of Ad5, with modifications to contain a carboxy-terminal decalysine tail for binding anionic cargo, including nucleic acids. The structure preserves the pentamerization ability of the penton base, enabling the formation of pinwheel structures. Exposure to cargo triggers the formation of the HPK bioparticle. Figures are reproduced from Sims et al (2018) J Controlled Rel, Alonso-Valenteen et al (2019) NAR, and Alonso-Valenteen et al (2025) Nat Nanotech with permission through the Creative Commons License.
Mechanism of endosomal lysis revealed by HPK
5. Cargo-induced nano-capsid assembly
A key insight from HPK research is that penton base capsomeres possess intrinsic self-assembly behavior. In isolation, capsomeres repel one another due to positive surface charge. Binding of anionic nucleic acids to the K10 domain neutralizes this charge, enabling cargo-templated assembly into polyhedral nano-bioparticles (NBPs) (Fig. 3).27, 30
These NBPs resemble viral capsids in size (20–40 nm), multivalency, and stability, protecting cargo from serum degradation while enhancing receptor avidity.27, 30
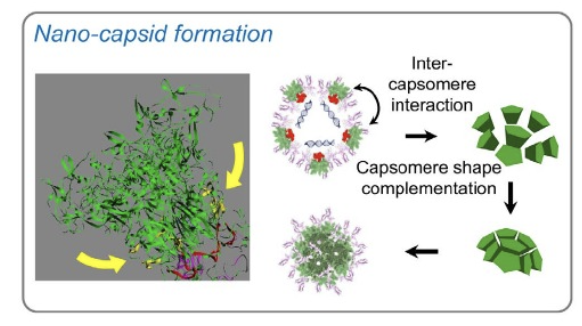
Fig. 3. HPK self-assembly. Ribbon structure of penton base capsomere portion of HPK highlighting the inter-capsomere binding domains (yellow arrows) that are enabled when cargo binding allows the capsomeres to converge and interact. The cork-shaped structure nucleates a polyhedral formation, encapsulating cargo in spherical particles. Figures are reproduced from Alonso-Valenteen et al (2019) NAR and Alonso-Valenteen et al (2025) Nat Nanotech with permission through the Creative Commons License.
6. Molecular mechanism of endosomal lysis
The most significant mechanistic advance from HPK studies is the elucidation of pH-triggered endosomal lysis.
6.1 Pentamer Destabilization
Molecular dynamics simulations and biophysical analyses revealed that penton base pentamers contain clusters of pH-sensitive residues, particularly histidines, lining the interior of the barrel-like capsomere (Fig. 4). 27, 30
Upon endosomal acidification (pH ~6.0–6.5), protonation of these residues induces electrostatic repulsion, driving pentamer dissociation.
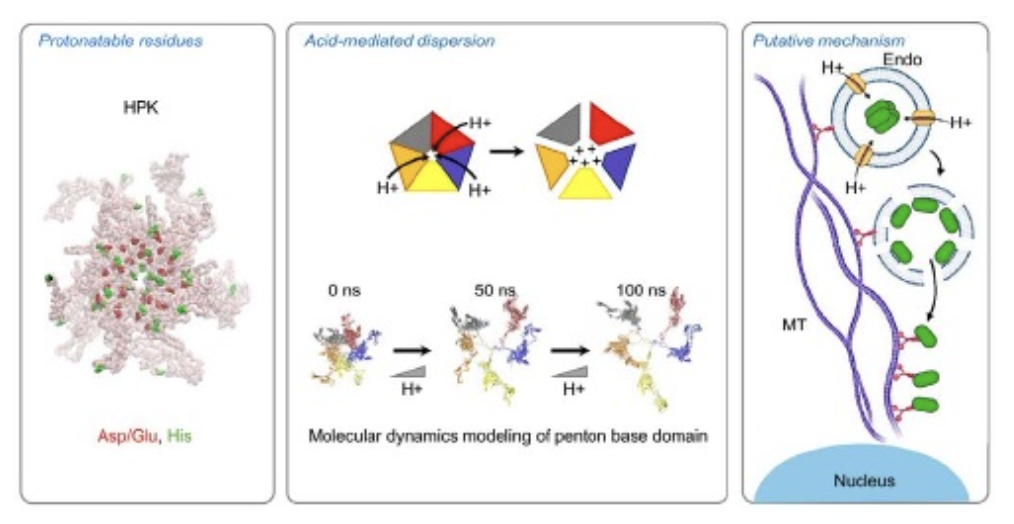 Fig. 4. Endosomolytic mechanism. Charged residues and histidines lining the solvent-exposed barrel of the HPK capsomere become protonated under increasingly acidic conditions, causing monomers to dissociate from one another by charge repellence, exposing hydrophobic domains that are likely to interact with endosomal membrane lipids and destabilize the endosomal membrane, allowing the escape of lumenal contents into the cytoplasm. Figures are reproduced from Alonso-Valenteen et al (2019) NAR with permission through the Creative Commons License.
Fig. 4. Endosomolytic mechanism. Charged residues and histidines lining the solvent-exposed barrel of the HPK capsomere become protonated under increasingly acidic conditions, causing monomers to dissociate from one another by charge repellence, exposing hydrophobic domains that are likely to interact with endosomal membrane lipids and destabilize the endosomal membrane, allowing the escape of lumenal contents into the cytoplasm. Figures are reproduced from Alonso-Valenteen et al (2019) NAR with permission through the Creative Commons License.
6.2 Membrane interaction
Pentamer dissociation exposes hydrophobic domains previously buried at monomer interfaces (Fig. 4).5, 27 These domains are likely to interact with endosomal lipids, causing direct membrane destabilization, rather than osmotic rupture.
Pharmacologic inhibition of endosomal acidification blocks HPK-mediated cytosolic delivery, confirming the pH-dependent mechanism.27, 30
6.3 Early endosomal escape
Intracellular trafficking and subcellular fractionation studies suggest that HPK escapes from early endosomes, avoiding lysosomal degradation and enhancing cytosolic bioavailability.27, 30
In vivo translation and broader implications
7. Cytosolic delivery of nucleic acids
HPK nano-capsids efficiently deliver siRNA and oligonucleotides into the cytosol,27-28, 31 achieving gene silencing and antitumor activity in vivo. Importantly, HPK recognizes both human and murine HER3, enabling validation in immunocompetent models. 27, 30
8. Immunological and functional implications
Compared with intact adenovirus vectors, HPK exhibits reduced immunogenicity because it lacks the viral genome and most capsid proteins, features that normally contribute to immune recognition.
Assembly of HPK into pentameric capsomeres decreases recognition by anti penton base antibodies, and subsequent formation of nano capsids completely shields the penton base domain while preserving HER3 specificity, collectively producing a ligand mimicking protomer that evades immunodetection by anti-viral antibodies.27, 30
9. Conceptual impact
HPK research demonstrates that viral entry functions can be mechanistically dissected and redeployed. The discovery that penton base pentamers act as pH-responsive, membrane-lytic switches reframes adenovirus biology and provides a blueprint for protein-based delivery systems that overcome endosomal barriers without synthetic polymers or replicating viruses.
Conclusion
The adenovirus penton base is a paradigmatic example of how viral evolution encodes solutions to intracellular delivery. Through HerPBK10 / HPK, these solutions have been translated into a receptor-targeted, self-assembling, endosomolytic nano-capsid platform. The elucidation of its endosomal lysis mechanism – pH-triggered pentamer dissociation and hydrophobic membrane interaction – bridges fundamental virology and therapeutic engineering, positioning the penton base as a cornerstone scaffold for next-generation delivery technologies.
References
Allen, R., Yokota, T., Endosomal Escape and Nuclear Localization: Critical Barriers for Therapeutic Nucleic Acids. Molecules 2024, 29 (24). PMC11677605
Staring, J., Raaben, M., Brummelkamp, T. R., Viral escape from endosomes and host detection at a glance. J Cell Sci 2018, 131 (15).
Russell, W. C., Adenoviruses: update on structure and function. J Gen Virol 2009, 90 (Pt 1), 1-20.
Nemerow, G. R., Stewart, P. L., Reddy, V. S., Structure of human adenovirus. Curr Opin Virol 2012, 2 (2), 115-21. PMC3322384
Zubieta, C., Schoehn, G., Chroboczek, J., Cusack, S., The structure of the human adenovirus 2 penton. Mol Cell. 2005, 17 (1), 121-35.
Reddy, V. S., Natchiar, S. K., Stewart, P. L., Nemerow, G. R., Crystal Structure of Human Adenovirus at 3.5 Å Resolution. Science 2010, 329 (5995), 1071-1075.
Liu, H., Jin, L., Koh, S. B. S., Atanasov, I., Schein, S., Wu, L., Zhou, Z. H., Atomic Structure of Human Adenovirus by Cryo-EM Reveals Interactions Among Protein Networks. Science 2010, 329 (5995), 1038-1043.
Medina-Kauwe, L. K., Endocytosis of adenovirus and adenovirus capsid proteins. Advanced Drug Delivery Reviews 2003, 55 (11), 1485-1496.
Bergelson, J. M., Cunningham, J. A., Droguett, G., Kurt-Jones, E. A., Krithivas, A., Hong, J. S., Horwitz, M. S., Crowell, R. L., Finberg, R. W., Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275 (5304), 1320-3.
Gaggar, A., Shayakhmetov, D. M., Lieber, A., CD46 is a cellular receptor for group B adenoviruses. Nat Med 2003, 9 (11), 1408-12.
Wang, H., Li, Z. Y., Liu, Y., Persson, J., Beyer, I., Moller, T., Koyuncu, D., Drescher, M. R., Strauss, R., Zhang, X. B., Wahl, J. K., 3rd, Urban, N., Drescher, C., Hemminki, A., Fender, P., Lieber, A., Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat Med 2011, 17 (1), 96-104.
Wickham, T. J., Mathias, P., Cheresh, D. A., Nemerow, G. R., Integrins avb3 and avb5 promote adenovirus internalization but not virus attachment. Cell 1993, 73 (2), 309-19.
Bai, M., Campis, L., Freimuth, P., Vitronectin receptor antibodies inhibit infection of HeLa and A549 cells by adenovirus type 12 but not by adenovirus type 2. Journal of Virology 1994, 68, 5925 – 5932.
Li, E., Stupack, D., Klemke, R., Cheresh, D. A., Nemerow, G. R., Adenovirus endocytosis via alpha(v) integrins requires phosphoinositide-3-OH kinase. J Virol 1998, 72 (3), 2055-61. PMC109499
Nakano, M. Y., Boucke, K., Suomalainen, M., Stidwill, R. P., Greber, U. F., The first step of adenovirus type 2 disassembly occurs at the cell surface, independently of endocytosis and escape to the cytosol. Journal of Virology. 2000, 74 (15), 7085-95.
Greber, U. F., Virus assembly and disassembly: the adenovirus cysteine protease as a trigger factor. Rev. Med. Virol. 1998, 8, 213-222.
Greber, U. F., Willetts, M., Webster, P., Helenius, A., Stepwise dismantling of adenovirus 2 during entry into cells. Cell 1993, 75 (3), 477-86.
Wiethoff, C. M., Wodrich, H., Gerace, L., Nemerow, G. R., Adenovirus protein VI mediates membrane disruption following capsid disassembly. J Virol 2005, 79 (4), 1992-2000.
Medina-Kauwe, L. K., Kasahara, N., Kedes, L., 3PO, a novel nonviral gene delivery system using engineered Ad5 penton proteins. Gene Therapy 2001, 8 (10), 795-803.
Rentsendorj, A., Xie, J., MacVeigh, M., Agadjanian, H., Bass, S., Kim, D. H., Rossi, J., Hamm-Alvarez, S. F., Medina-Kauwe, L. K., Typical and atypical trafficking pathways of Ad5 penton base recombinant protein: implications for gene transfer. Gene therapy 2006, 13 (10), 821-836.
Hong, S. S., Gay, B., Karayan, L., Dabauvalle, M. C., Boulanger, P., Cellular uptake and nuclear delivery of recombinant adenovirus penton base. Virology. 1999, 262 (1), 163-77.
Fender, P., Ruigrok, R. W., Gout, E., Buffet, S., Chroboczek, J., Adenovirus dodecahedron, a new vector for human gene transfer [see comments]. Nature Biotechnology 1997, 15 (1), 52-6.
Seth, P., Willingham, M. C., Pastan, I., Binding of adenovirus and its external proteins to Triton X-114. Dependence on pH. Journal of Biological Chemistry. 1985, 260 (27), 14431-4.
Seth, P., Fitzgerald, D., Ginsberg, H., Willingham, M., Pastan, I., Evidence that the penton base of adenovirus is involved in potentiation of toxicity of Pseudomonas exotoxin conjugated to epidermal growth factor. Molecular & Cellular Biology 1984, 4 (8), 1528-33.
Medina-Kauwe, L. K., Maguire, M., Kasahara, N., Kedes, L., Nonviral gene delivery to human breast cancer cells by targeted Ad5 penton proteins. Gene Ther 2001, 8 (23), 1753-61.
Benhaghnazar, R. L., Medina-Kauwe, L., Adenovirus-Derived Nano-Capsid Platforms for Targeted Delivery and Penetration of Macromolecules into Resistant and Metastatic Tumors. Cancers 2023, 15 (12). PMC10296971
Alonso-Valenteen, F., Pacheco, S., Srinivas, D., Rentsendorj, A., Chu, D., Lubow, J., Sims, J., Miao, T., Mikhael, S., Hwang, J. Y., Abrol, R., Medina Kauwe, L. K., HER3-targeted protein chimera forms endosomolytic capsomeres and self-assembles into stealth nucleocapsids for systemic tumor homing of RNA interference in vivo. Nucleic acids research 2019, 47 (21), 11020-11043. PMC6868389
Sims, J. D., Taguiam, J. M., Alonso-Valenteen, F., Markman, J., Agadjanian, H., Chu, D., Lubow, J., Abrol, R., Srinivas, D., Jain, A., Han, B., Qu, Y., Mirzadehgan, P., Hwang, J.-Y., Rentsendorj, A., Chung, A., Lester, J., Karlan, B. Y., Gray, H. B., Gross, Z., Giuliano, A., Cui, X., Medina-Kauwe, L. K., Resistance to receptor-blocking therapies primes tumors as targets for HER3-homing nanobiologics. Journal of Controlled Release 2018, 271, 127-138.
Kilroy, M. K., Park, S., Feroz, W., Patel, H., Mishra, R., Alanazi, S., Garrett, J. T., HER3 Alterations in Cancer and Potential Clinical Implications. Cancers 2022, 14 (24). PMC9776947
Alonso-Valenteen, F., Mikhael, S., Wang, H., Sims, J., Taguiam, M., Teh, J., Sances, S., Wong, M., Miao, T., Srinivas, D., Gonzalez-Almeyda, N., Cho, R. H., Sanchez, R., Nguyenle, K., Serrano, E., Ondatje, B., Benhaghnazar, R. L., Gray, H. B., Gross, Z., Yu, J., Svendsen, C. N., Abrol, R., Medina-Kauwe, L. K., Systemic HER3 ligand-mimicking nanobioparticles enter the brain and reduce intracranial tumour growth. Nat Nanotechnol 2025, 20 (5), 683-696. PMC12095042
Agadjanian, H., Chu, D., Hwang, J. Y., Wachsmann-Hogiu, S., Rentsendorj, A., Song, L., Valluripalli, V., Lubow, J., Ma, J., Sharifi, B., Farkas, D. L., Medina-Kauwe, L. K., Chemotherapy targeting by DNA capture in viral protein particles. Nanomedicine (London, England) 2012, 7 (3), 335-52. PMC3429352
Stewart, P. L., Chiu, C. Y., Huang, S., Muir, T., Zhao, Y., Chait, B., Mathias, P., Nemerow, G. R., Cryo-EM visualization of an exposed RGD epitope on adenovirus that escapes antibody neutralization. EMBO Journal. 1997, 16 (6), 1189-98.