
Disentangling the processes that drive the evolution of complex biological structures is a major aim of evolutionary biology. Multivariate data in comparative studies can reflect various signals (e.g., ecological, allometric, phylogenetic, or combinations thereof; [32]). In this study, we employed 3D landmark configurations, a functional subdivision of the vertebral column in 24 dolphin species (Family Delphinidae), and phylogenetic comparative methods to test the effects of phylogenetic, ecological and allometric signals along the vertebral column of the most diverse cetacean family.
Our results indicate that vertebral morphology in dolphins is shaped by distinct but overlapping signals: strong ecological effects on the thorax-torso boundary (ThTo), the mid torso (Tm), and the synclinal point (SP); vertebral size effects, particularly ThTo and SP; and phylogenetic constraints on Tm, SP, and the tailstock (TS). These findings suggest that different regions of the column evolve under varying combinations of ecological, allometric, and phylogenetic influences.
Ecological signal
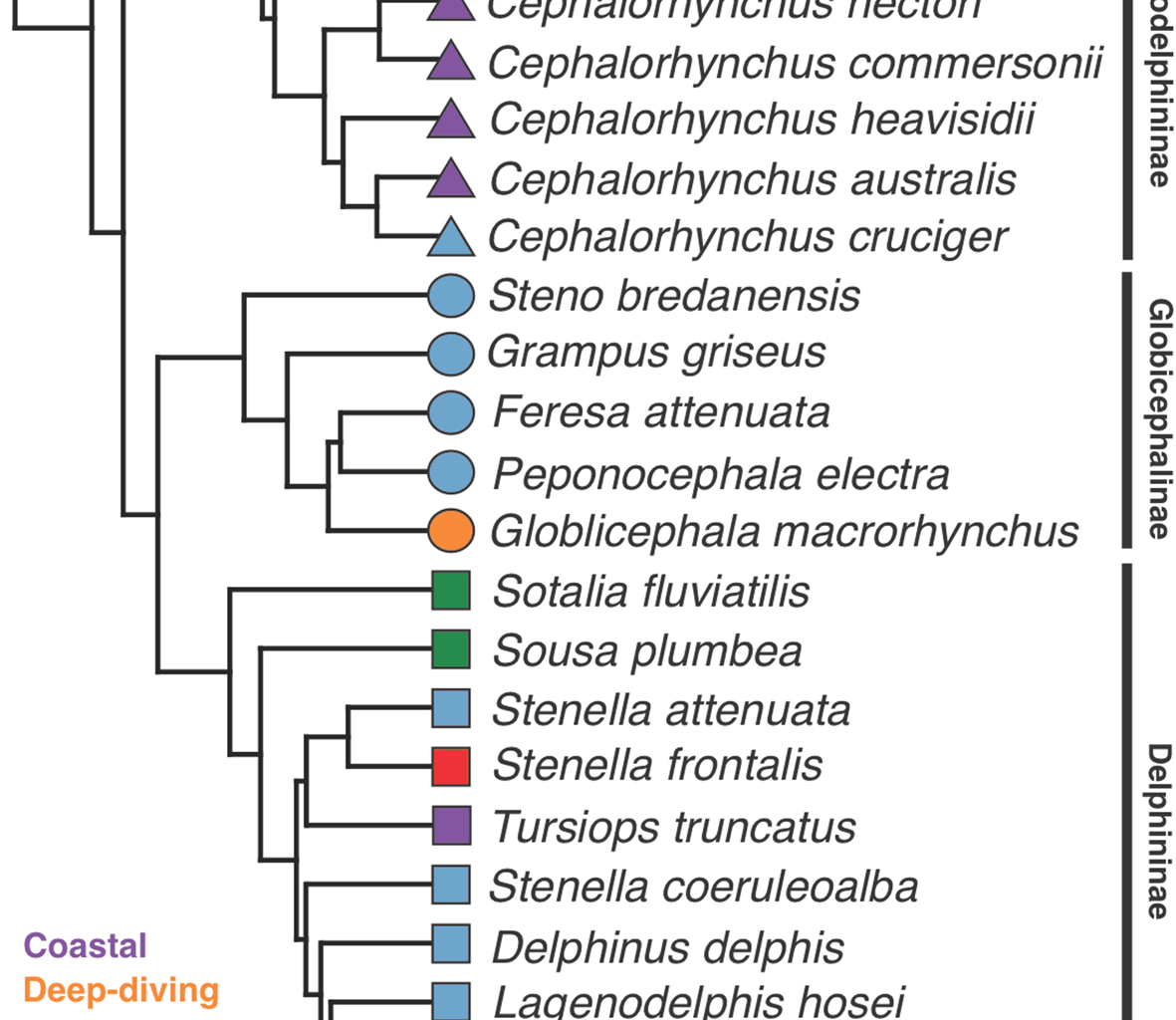
Previous studies on cetacean vertebral morphology have mainly focused on ecological signal [8,9,10, 14, 15, 25]. Based on vertebral morphology, it has been proposed that the most recent common ancestor (MRCA) of crown delphinids, as well as those of each subfamily (Lissodelphininae, Globicephalinae, and Delphininae), likely inhabited offshore environments, showing morphologies associated with oceanic non-fast-swimming species, with subsequent shape diversification linked to the biomechanical demands of different habitats and habits [10, 11]. Our results revealed ecological signal in three regions critical for swimming efficiency: ThTo, Tm, and SP.
Both the anterior and middle torso (ThTo and Tm) are especially important regions, as they are where the longissimus muscle generates the greatest forces that are transferred to the flukes [42]. In these regions, habitat explained more than 37% of total variation, with relatively high Z values, suggesting that habitat has a greater effect on vertebral morphology than that expected by chance. The synclinal point marks the transition between the stable torso and the flexible tailstock, representing the area where muscle forces that affect the fluke’s angle of attack are produced [15, 25, 43, 44]. Here, the effect of habitat was smaller than in the first two regions, with relatively low R2 but high Z values. The associations between shape and habitat in these three areas suggest a particular influence of biomechanical constraints on the vertebral morphology imposed by habitat differences. Similarly, the distribution of shape in phylomorphospace when the ecological signal was maximised resembled the patterns described by Marchesi [11], with closely related species showing distinct morphologies associated with particular habitats. Species with specific habitat requirements (e.g., deep diving, rivers and bays, or extremely fast-swimming species) showed marked divergence from their closest ancestors (see also [11]).
Importantly, the classification of habitats into discrete categories is inherently subjective and does not reflect the continuum of delphinid habitats. Other ecological factors (feeding mechanism, migration, prey size, etc.) may also influence vertebral morphology. For example, burst swimming speed in cetaceans has been linked to vertebral morphology and the vertebral count, with consequences for vertebral regionalisation [14]. In our study, differences in phylomorphospace occupation among species with similar habitats may reflect fine-scale ecological partitioning between sympatric species, as morphology could be linked to differences in prey size or spatial distribution [45,46,47]. While ecological drivers explained much of the shape variation in these regions, size effects also contributed, as discussed below.
Allometric effects
Evolutionary allometry has been shown for the skulls, mandibles, and vertebrae of cetaceans [21,22,23,24]. For the odontocete skull, Vicari et al. [23] reported that families occupy distinct positions via allometric regression, with 9% of skull shape variation explained by size. With respect to vertebral morphology, Marchesi et al. [22] reported strong size-shaped associations across most regions of the column in Lissodelphininae dolphins, suggesting that size changes may have facilitated rapid morphological diversification within a short evolutionary timeframe.
In our study, size significantly influenced vertebral shape after accounting for phylogeny in ThTo and, to a lesser degree, in SP, possibly reflecting rapid evolutionary changes mediated by size in biomechanically critical regions. This was particularly evident in species with habitat requirements differing from those of the delphinid common ancestor, such as the large deep diver Globicephala macrorhynchus, the large riverine Sousa plumbea, and the small coastal Cephalorhynchus (within Lissodelphininae), for which a reduction in size has already been proposed as a driver of morphological change [48, 11]. Importantly, the influence of size on vertebral shape may differ across regions and have different biomechanical implications [22]. These results should be interpreted with caution, as our sample may be biased regarding sex, and sexual size dimorphism is known in delphinids [49].
Phylogenetic signal
Studies on phylogenetic signals in cetaceans remain scarce. Vicari et al. [23] reported phylogenetic signals for both skull size and shape, which were stronger for size (Kmult = 0.653) than for shape (Kmult = 0.565), with size linked to ecological traits but shape showing no association. Conversely, Galatius et al. [21] reported strong phylogenetic signals in delphinid skull shapes, with subfamilies displaying distinct morphologies. For vertebrae, Viglino et al. [16] detected phylogenetic signals in traits related to swimming muscle architecture across delphinids and Pontoporia blainvillei, but this signal disappeared when Pontoporia was excluded, suggesting a strong effect of this distantly related species and highlighting the importance of considering several species in this type of study. Marchesi et al. [50] reported no significant phylogenetic signal within porpoises, likely due to parallel convergence, but strong signals were recovered across delphinoids, particularly in Tm, suggesting supra-family constraints during delphinoid diversification (~ 20 Mya [50]).
Our results parallel these findings: a phylogenetic signal was detected along the dolphin vertebral column in centroid size (CS), shape (Procrustes coordinates), and ordination methods maximising ecological (PhyPCA) or phylogenetic (PACA) signals. Size exhibited a weak but significant phylogenetic signal (Kmult < 0.62, Z < 2.07) in the three most posterior regions, which are considered sub regions within the caudal region [14], suggesting that partial phylogenetic constraints on vertebral size in these areas are responsible for differences in swimming performance. With respect to shape (Procrustes coordinates, PhyPCA, and PACA), the phylogenetic signal was evident to varying degrees in all regions except ThTo (in PhyPCA). Disregarding the dataset,, with the lowest Kmult values occurred in the anterior regions, while higher values were found for Tm (Kmult ≅ 0.8) and TS (Kmult ≅ 0.9). The values for SP were intermediate, although the Z values were the highest for this region. This pattern suggests that shape diversification may have contributed significantly to delphinid radiation, with phylogeny constraining shape in biomechanically relevant regions.
Consistent with earlier studies [16, 22], phylogenetic signals in these regions may be stronger with broader taxonomic sampling, implying constraints at the family level that were weakened during subfamily radiation. The strong phylogenetic signal in Tm and TS supports the idea that subfamily level constraints shape vertebral morphology during early delphinid diversification (12–6 Mya [26],).
Interaction of signals
A low phylogenetic signal in Th and ThTo for which habitat and size showed significant effects with an important percentage of shape variance explained by each factor suggest the combination of multiple signals at various degrees along the vertebral column. Importantly, the coexistence of strong phylogenetic and ecological signals in Tm and SP, with marked shape differences among closely related species in contrasting habitats, suggests that multiple interacting factors shape vertebral morphology to varying degrees depending on the level of the analysis (family or subfamily). In this way, within each subfamily, we observed highly divergent shapes that were related to the biomechanical demands of each habitat and may have been involved in the explosive radiation within subfamilies in the late Miocene–Pliocene (5–2 Mya; [3, 4, 26]. Similarly, Vicari et al. [24] reported a strong phylogenetic signal with a jaw shape (Kmult ≅ 1.1) alongside a significant association with the bio-sonar mode and diet, indicating an ecological signal in conjunction with a phylogenetic signal in an ecologically relevant structure.
Dimensionality and hierarchical processes
Phenotypic evolution is widely recognised as a multivariate process [40, 41, 51, 52], where complex traits such as shape evolve in correlated dimensions. In this sense, phylogenetic signals may also be separated into multiple dimensions in the data [32, 40]. Here, Kmult values below 1 in all cases, the presence of both ecological and phylogenetic signals, and the results of the ordination analyses collectively suggest that multiple signals (ecologic, allometric, and phylogenetic) influence vertebral morphology. Nevertheless, our models assumed Brownian motion, which may be inadequate for clades with rapid diversification [53]. Within this evolutionary framework, vertebral morphology modification patterns seem to play a key role, suggesting continuity between evolutionary processes occurring at different phylogenetic levels and rapid ecomorphological changes at the macroevolutionary level [10]. Our results suggest that the morphology of Tm and TS, and to a lesser extent SP, was constrained by subfamily level divergence, whereas the morphology of Th and ThTo diverged from that of the MRCA, possibly due to ecological factors.
The vertebral column is a modular structure subject to hierarchical developmental processes, producing phenotypic units capable of independent modification [54]. Our findings demonstrate that different evolutionary signals might differentially affect the vertebral morphology of dolphins depending on the region along the skeleton, phylogenetic level, and shape dimension. At the family level, the vertebral morphology of Th and ThTo is partially constrained by phylogeny but, more importantly, by other factors, such as ecology or size. In contrast, the vertebral morphology of Tm, SP and TS was mostly constrained by phylogeny. Nonetheless, ecological signals are also important in critical regions for the biomechanics of fast-swimming oceanic species (ThTo, Tm, and SP), highlighting the role of ecological factors at relatively fine scales [9, 11, 22].
Methodological implications and future directions
Further work applying evolutionary models to identify the best-fit evolutionary scenarios for Delphinidae is needed to fully characterize evolutionary rates, selection strengths, and constraints, and to determine the extent to which additional ecological signals could explain vertebral morphological diversification. In particular, OU-based approaches could test whether different regions of the vertebral column have been shaped by attraction toward adaptive optima, while convergence metrics [55, 56] could be used to evaluate whether taxa occupying similar habitats (e.g., coastal versus offshore environments) have independently evolved similar vertebral shapes.
However, current methodological frameworks impose important constraints for such analyses when working with high-dimensional landmark data. Most implementations of evolutionary models (e.g., [57,58,59,60]) are designed for univariate traits, while packages that accommodate multivariate data (e.g., [55, 61]) are limited to datasets with relatively few dimensions. Applying these methods to landmark-based datasets typically requires dimensionality reduction (usually via phylogenetic principal component analysis) as was employed by different authors (e.g., [62,63,64]), which can obscure evolutionary trends of shape variation and potentially bias evolutionary inference [65, 66]. For this reason, we adopted a Brownian motion framework, which remains the most common and useful model for investigating macroevolutionary patterns in high-dimensional morphometric datasets [40] without strong assumptions about adaptive optima.
Future studies could combine dimensionality-reduction approaches with OU-based models to begin testing explicit adaptive hypotheses, but ideally, methodological developments that allow for parameter estimation directly from high-dimensional data will be needed to rigorously evaluate habitat-driven constraints and convergence in vertebral morphology. Furthermore, with the recent development of methods incorporating intraspecific variation into PGLS analyses (Adams & Collyer 2024), future research should also explore how accounting for within-species variability affects estimates of ecological and phylogenetic signals. This is particularly relevant for Delphinidae, where intraspecific variation in vertebral number and shape may influence functional interpretations. Thus, these future directions will allow a more powerful evaluation of how ecological pressures have shaped the vertebral column, complementing the methodology provided here on the distribution of phylogenetic, ecological, and allometric influences across different regions.