
Of the pectoral musculature, we have selected the five muscles (i.e., mm. deltoideus scapularis/major, pectoralis, supracoracoideus, coracobrachialis brevis p. ventralis/p. cranialis, biceps brachii) due to the fact that they are the largest and perform the main movements of the forelimb. The origins and insertions of these pectoral muscles are studied in different groups of extant archosaurs (crocodiles and birds) in order to hypothesize their functions in four selected taxa of basal paravians (i.e., Bambiraptor, Buitreraptor, Archaeopteryx, and Confuciusornis).
M. deltoideus scapularis/major
In living crocodiles, the m. deltoideus scapularis originates in the distal end of the lateral surface of the scapular blade [18, 43]. This muscle inserts on the dorsal surface of the proximal end of the humerus close to the humeral head. In these archosaurians, the function of this muscle is to abduct the humerus [18], Table 1).
Table 1 Function of five pectoral girdle muscles of different archosaur groups (e.g., crocodile, paravians, basal birds, non-volant paleognath and neognath). Abbreviations: BB, m. biceps brachii; CBB/CBCr, m. coracobrachialis brevis p. ventralis/p. cranialis; DM/DS, m. deltoideus scapularis/major; P, m. pectoralis; SC, m. supracoracoideus
In living birds, the m. deltoideus scapularis is homologous with the m. deltoideus major [18, 36]. In neognath birds and Tinamiformes, the origin of this muscle is on the lateral surface of the scapula, in the medial border of the acromion [18, 35, 36]. The insertion is into the caudal edge of the deltopectoral crest, and its function is to retract and elevate the humerus [18], Table 1). Particularly in Rhea and other non-volant paleognaths, the origin is also established on the lateral aspect of the proximal end of the scapula and inserts widely, extending beyond the middle of the dorsal border of the humerus [18, 20].
Due to the similarity of the scapula blade among non-avian paravians (e.g., Bambiraptor, Buitreraptor), basal birds (e.g., Archaeopteryx, Confuciusornis), and extant birds [2, 11, 12, 15], we infer that the m. deltoideus scapularis/major would be also similar in all these living and extinct paravians. The origin could reach the medial border of the proximal end of the scapula, and could extend over the lateral surface of the scapula, towards the glenoid cavity, occupying a larger area than that found in extant flying birds, as is observed in non-volant paleognaths (see [20]).
Regarding the insertion of the m. deltoideus scapularis/major, it could be anchored on the first section of the deltopectoral crest, on its caudal side, as occurs in living crocodiles [18, 43] and extant birds [18, 20, 30].
As suggested by Olson & Feduccia [44] and Ruben [45], the m. deltoideus major rather than m. supracoracoideus may have been the main elevator of the humerus in Archaeopteryx. We concur with these authors’ proposal, considering that it origin is located slightly dorsal and medially to the glenoid cavity (Fig. 2) generating an elevator function of the humerus. The same function may be also applied to the m. deltoideus scapularis/major of Bambiraptor, Buitreraptor, and Confuciusornis, as well (Table 1). In this way, the m. deltoideus scapularis/major together with other elevator muscles present in extant diapsids (i.e., the m. latissimus dorsi group; [20, 43, 46]), would have performed a fundamental role as the exclusive elevators of the humerus in paravians and basal birds. With the morphological changes occurring in the scapular girdle of the derived birds (i.e., acrocoracoid well developed anterodorsally, glenoid cavity subhorizontal oriented, scapulocoracoid articulation located caudally from glenoid cavity in Enantiornithes, and Euornithes), the function of main elevator of humerus came to be carried out mainly by the m. supracoracoideus which is primarily responsible for the rapid upstroke of the wing in extant volant birds [41, 47]. We would expect the morphological transition between these taxa to have been gradual, but the fossil evidence known to date suggests that there would have been abrupt osteological changes in the pectoral girdle and forelimbs between Confuciusornithidae and Enantiornithes (see also [10, 11]).
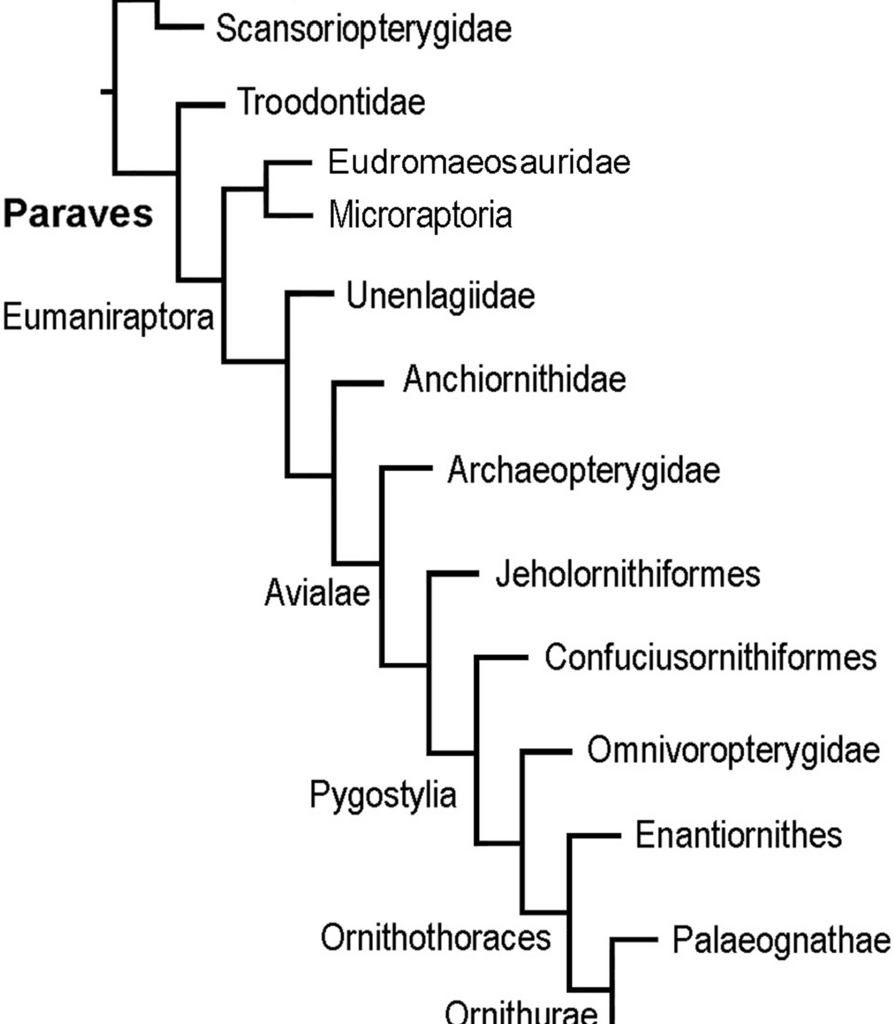
Fig. 2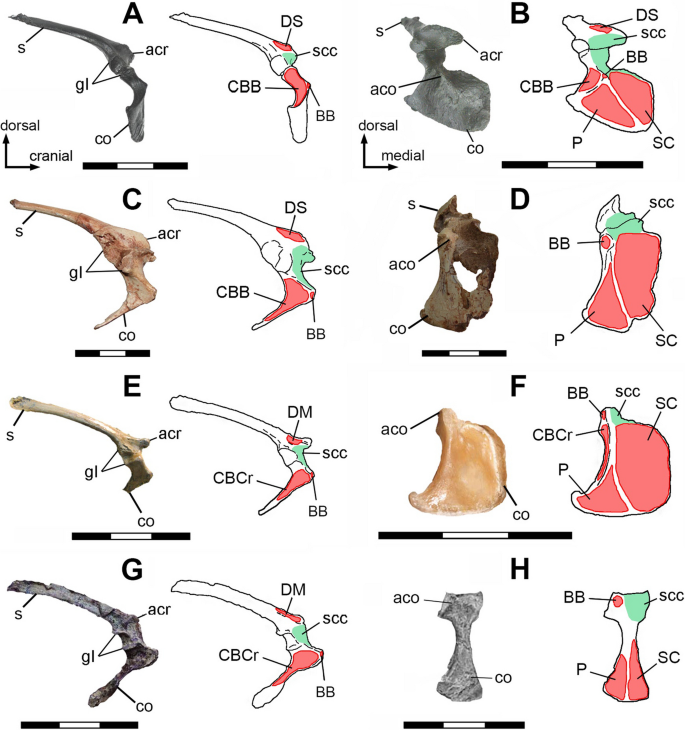
Muscular maps showing the sites of origin (red) of right scapula and coracoid of Bambiraptor feinbergi AMNH 001 (A, B); Buitreraptor gonzalezorum MPCN-PV-598 (C, D); Archaeopteryx lithographica WDC-CSG-100 (E rotated and modified from [48], F); Confuciusornis sanctus BMNHC PH766 (G, rotated and modified from [49]), and Confuciusornis sanctus GMV-2132 (H; modified from [50]), in lateral (A, C, E, G) and cranial (B, D, F, H) views. Abbreviations: aco, acrocoracoid process; acr, acromion, co, coracoid; gl, glenoid cavity; s, scapula, scc, supracoracoidal canal; BB, m. biceps brachii; CBB, m. coracobrachialis brevis p. ventralis, CBCr, m. coracobrachialis p. cranialis; DM, m. deltoideus major; DS, m. deltoideus scapularis; P, m. pectoralis; SC, m. supracoracoideus. Light green, supracoracoidal canal surface. Scale bars 3 cm
M. pectoralis
In all present-day archosaurs (crocodiles and birds), the m. pectoralis is composed of one to three bellies which originate at different parts of the pectoral girdle: ventral to the glenoid cavity, at the coracoid and/or on the sternum; in some cases, the fibers of the m. pectoralis reach the furcula (i.e., Tinamiformes and Neognathae), or are attached over the sternal ribs (as in non-avian reptiles and neognaths; [18, 30, 36, 43]). In all cases, the m. pectoralis inserts onto the cranial aspect of the proximal end of humerus, at the dorsal edge of the deltopectoral crest [18, 20, 23, 30, 36, 43].
Its function in crocodiles is to adduct the humerus [43], Table 1). In Tinamiformes and neognaths, the main function of the m. pectoralis is to depress the humerus, although it also assists in its adduction [18, 41], Table 1). In non-volant paleognaths the function of m. pectoralis is different from other extant birds, its main role being protracting and depressing the humerus [18], because its origin is ventral and cranioventral with respect to its insertion and the glenoid cavity [20], Table 1).
Based on the coracoid attachments observed in modern flying birds and particularly in Rhea, it is inferred that, in the selected fossil taxa, the m. pectoralis also originated on the cranial side of the coracoid [10], on its lateral half, ventral to the glenoid cavity and the acrocoracoid process (Fig. 2). It is possible that its origin also expanded over the gastralia or sternal plates (bony or cartilaginous), even when the sternal keel was not well-developed. In turn, it can be inferred that it could even cover part of the sternal ribs and reach the lateral border of the furcula and the sternocoracoclavicularis membrane [10, 42, 44].
In Bambiraptor, Buitreraptor, Archaeopteryx, and Confuciusornis it is inferred that the insertion of the m. pectoralis was located on the craniodorsal margin of the deltopectoral crest of the humerus (Fig. 3; [18, 23]), as occurs in reptiles and modern birds [18, 20, 23, 30, 36, 41, 43].
Fig. 3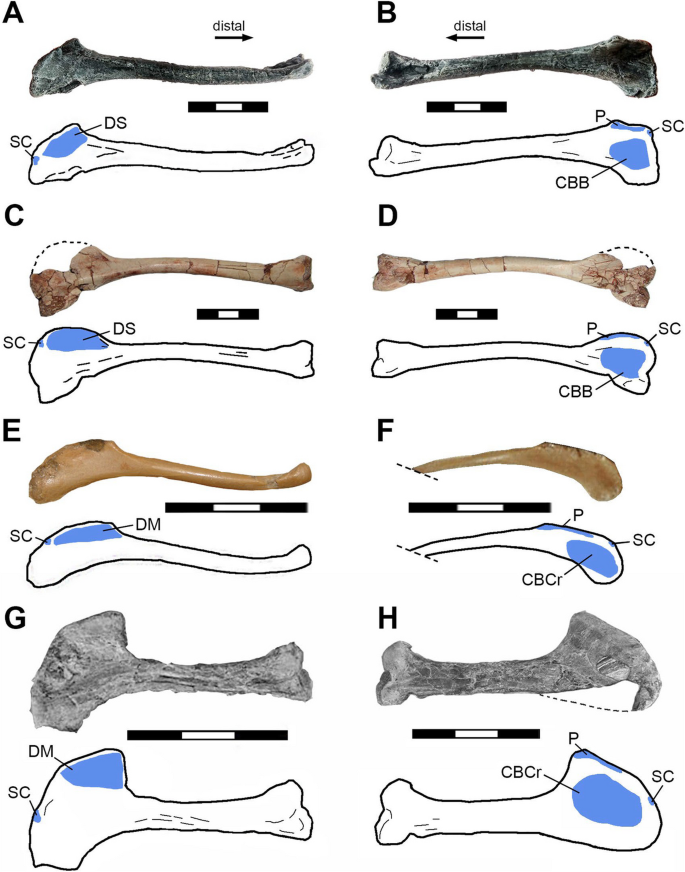
Muscular maps showing the sites of insertion (blue) of right humerus of Bambiraptor feinbergi AMNH 001 (A, B); Buitreraptor gonzalezorum MPCA 245 (C, D); Archaeopteryx lithographica MB.Av.101 (E); Archaeopteryx lithographica WDC-CSG-100 (F; the distal end is not uncovered); Confuciusornis sanctus GMV-2133 (G*; rotated and modified from [50]),and Confuciusornis sanctus GMV-2132 (H*; rotated and modified from [50]),in caudal (A, C, E, G) and cranial (B, D, F, H) views. Abbreviations: CBB, m. coracobrachialis brevis p. ventralis, CBCr, m. coracobrachialis p. cranialis; DM, m. deltoideus major; DS, m. deltoideus scapularis; P, m. pectoralis; SC, m. supracoracoideus. Scale bars 3 cm
Although the coracoid of Bambiraptor, Buitreraptor, Archaeopteryx, and Confuciusornis lacks bony scars indicating a separation between the origin of the m. pectoralis and the m. supracoracoideus, it can be thought that they bordered each other, but did not overlap. The growth in volume of the m. pectoralis, like the overlapping of the m. supracoracoideus, can be inferred to have occurred in more derived taxa of Avialae, where there was a great development of the sternal keel [10, 51], and consequently the m. pectoralis volume has been greater reaching the furcula.
Considering that the main protractor function of the m. pectoralis in extant crocodilians and birds, it is suggested that in Bambiraptor, Buitreraptor, Archaeopteryx, and Confuciusornis the function was similar and protracted the humerus in a cranioventral direction and adducted the arm (Table 1), in agreement with what was proposed by Ostrom [1] and Novas et al. [11]. This can be explained based on the ventral, ventrocaudal and ventrocranial location of the origin of this muscle (i.e., on the coracoid, sternum and/or furcula, respectively) with respect to the glenoid cavity. In all cases, this muscle would have been inserted directly in the humerus, without generating any type of pulley with the acrocoracoid process.
M. supracoracoideus
In non-avian reptiles the m. supracoracoideus is made up to two or three bellies, which can originate mainly on the scapula and coracoid, positioning itself cranially to the glenoid cavity (e.g., Chamaeleo, Alligator, Caiman; [23, 43, 52]). Although in paleognaths and neognaths the origin is ventral to the glenoid cavity, its anchorage area may be the coracoid and/or the sternum. In tinamids and neognaths the origin is wide, reaching the lateral and basal area of the sternal keel, the lateral margin of the furcula next to the sternocoracoclavicularis membrane, and the medial part of the coracoid and the processus procoracoideus [11, 18, 30, 31, 35, 36]. In non-flying paleognaths (e.g., Rhea, Struthio, Dromaius) the origin of the m. supracoracoideus is located on the cranial side of the coracoid, with a small surface area covering the sternum or without anchoring on it [18, 20, 31].
The insertion of this muscle in extant crocodilians and living birds is located on the dorsal-dorsocranial side of the proximal end of the humerus, usually at the proximal end of the deltopectoral crest [18, 23, 30, 36, 43]. The m. supracoracoideus group in non-avian reptiles (e.g., Crocrodylia; [43]), and flightless paleognaths (e.g., Rhea, Struthio, [11, 18, 20]) plays a role of humeral protractor. In neognaths and Tinamiformes this muscle is channeled into the “triosseal canal” (structure bounded by the acrocoracoid, acromion, and furcula), and its main function is to elevate and abduct the wing [22, 30, 35, 41, 53], Table 1).
Considering that the specimens of Buitreraptor gonzalezorum MPCN-PV-598, MPCA 245, Archaeopteryx lithographica MB.Av.101, WDC-CSG-100, and Confuciusornis sanctus BMNHC PH766, GMV-2132/2133, did not preserve an ossified sternum, and that Bambiraptor feinbergi AMNH 001 present two sternal plates but did not preserve an ossified keel, it is inferred that in these taxa the m. supracoracoideus had a smaller development than in extant volant birds [1, 10, 19, 45]. In the above mentioned extinct paravians the m. supracoracoideus could have originated on the cranial and medial surfaces of the coracoid, although it has probably extended caudally to reach the sternal plates or even the gastral ribs [10, 54].
Buitreraptor, Bambiraptor, Archaeopteryx and Confuciusornis do not present a “triosseal canal” because the furcula could contact the acromion but did not reach the acrocoracoid process [1, 12, 55, 56]. In these taxa, and non-volant paleognaths (i.e., Rhea, Struthio), a supracoracoidal canal is formed by the concave area between the acromion and acrocoracoid process [11, 20]. The presence of this canal in Buitreraptor, Bambiraptor, Archaeopteryx and Confuciusornis, suggests that the m. supracoracoideus was channeled through this space (Figs. 2; 4; see Additional file 1). In these taxa the supracoracoidal canal was probably tapered by the acrocoraco-acromial ligament that connects both processes as occurs in the extant Rhea [11, 20]. The insertion of the m. supracoracoideus would be located on the dorsal margin of the deltopectoral crest, being proximal to the insertion of the m. pectoralis (Figs. 3, 4).
Fig. 4
Three-dimensional model of the m. supracoracoideus (red) on the right pectoral girdle and humerus of Buitreraptor gonzalezorum (MPCN-PV-598) in dorsal (A) and cranial (B) views. Scale bar 3 cm
Based on the muscle anchoring sites inferred for Buitreraptor, Bambiraptor, Archaeopteryx and Confuciusornis, the origin of the m. supracoracoideus would be located ventromedially to the glenoid cavity (Figs. 2; 4; see Additional file 1), thus generating an obliquely oriented force (in cranial view), from a laterodorsal to medioventral direction, following the orientation of the supracoracoidal canal [11]. In the particular case of Archaeopteryx, some authors like Ostrom [1], Walker [57] and Olson & Feduccia [44], indicate that the shape of the coracoid would not allow the m. supracoracoideus to form a pulley, a conclusion that we agree. In our interpretation, the main probable function of the m. supracoracoideus in Buitreraptor, Bambiraptor, Archaeopteryx and Confuciusornis was the protraction of the humerus, in contrast to other authors that support an elevator function of this muscle (see Pittman et al., 2022). This protractor function would be more similar to what occurs in Rhea and other non-flying paleognaths than the elevator function performed by extant volant birds [20], see Table 1).
We agree with Mayr [10] in that the m. supracoracoideus of basal birds did not have a large origin as occurs in crown group birds. However, it is important to emphasize here some issues about the m. supracoracoideus origin that were previously overlooked. We propose that even assuming that this muscle had a major origin in paravians and basal birds (i.e., onto the furcula, sternocoracoclavicular membrane, gastralia, and/or sternum/sternal plates), it would still be routed through the supracoracoidal canal (see [11]). Therefore, the enlargement of the area of origin cranially, ventrally and/or caudally with respect to the glenoid cavity, would not modify the protraction function of the m. supracoracoideus. The same condition can be found in Enantiornithes as well, as the m. supracoracoideus passes through the supracoracoidal canal, in the medial side of the coracoid. Therefore, the elevating function of this muscle would not change if it were attached on the coracoid or distally on the sternum.
As Biewener & Roberts [58] have shown, an anchorage of m. supracoracoideus placed further away from the glenoid cavity would have generated a change in the power output employed by the muscle, being greater with a larger anchorage area and muscle mass.
Thus, with the information obtained in the present paper, we suggest that the key factors that have modified the function of the m. supracoracoideus include the type of scapulocoracoid joint, the inclination and orientation of the supracoracoidal canal, and the orientation of the glenoid cavity. A change in the origin of the muscle, from a cranial (i.e., on the coracoid) to a more caudal position (i.e., on the sternum), would not influence the route through the supracoracoidal canal.
M. coracobrachialis brevis p. ventralis/p. cranialis
In extant non-avian reptiles (i.e., Crocodylia) the m. coracobrachialis brevis p. ventralis originates on the lateral surface of the coracoid and close to its caudal edge, being ventral to the glenoid cavity [18, 43]. The insertion is located at the proximal end of the humerus, on its ventral side and close to the cranial edge [18, 43].
In non-volant paleognaths the m. coracobrachialis p. cranialis originates from the entire area of the subglenoid fossa and inserts on the cranial aspect of the proximal end of the humerus, between the deltopectoral crest and the bicipital crest [11, 18, 20, 23, 31]. In Tinamiformes and neognath birds the m. coracobrachialis p. cranialis originates at the end of the acrocoracoid process, being ventral to the origin of the m. biceps brachii, and its insertion occurs on the cranial aspect of the proximal end of the humerus, ventrally to the insertion of the m. pectoralis [41, 59].
The function of this muscle in non-avian reptiles is to protract and depress the humerus [18], thus generating adduction of the forelimb (Table 1). In non-volant paleognaths, like Rhea, its function is to depress the humerus [11, 18], Table 1), due to the ventral position of the origin with respect to the glenoid cavity. In contrast, in extant flying birds, this muscle has a protractor function [41], Table 1) because its origin is located in a craniodorsal position and is medial to the glenoid cavity.
The subglenoid fossa of Bambiraptor, Buitreraptor, Archaeopteryx, and Confuciusornis may represent the origin of the m. coracobrachialis brevis p. ventralis/p. cranialis (Fig. 2), and its insertion must have occupied the cranial area of the proximal end of the humerus, being ventral to the edge of the deltopectoral crest Fig. 3; [38]. Due to the ventral location of the origin of this muscle with respect to the glenoid cavity, it is considered that the function in all the selected taxa was twofold (Table 1): first, the protraction of the humerus would be performed when the forelimb was folded or close to the body [1, 18], and second, the depression of the humerus would occur when the forelimb was extended, in agreement with the proposal of Nicholls & Russell [23].
M. biceps brachii
In extant non-avian reptiles (i.e., Crocodylia) the m. biceps brachii originates from the cranial edge of the lateral surface of the coracoid, taking a cranioventral position to the glenoid cavity [18, 43]. The insertion in these extant archosaurs is found only at the proximal end of the radius [18, 43].
In neognath and paleognath birds, the main origin of the m. biceps brachii is located at the end of the acrocoracoid process [18, 36, 60, 61], whereas in some birds it originates on the lateral border of the coracoid (e.g., Rhea, [20, 62]), or on the ventral border of the cranial side of the proximal end of the humerus (e.g., Tyto furcata, [30, 36]). The insertion is mostly at the proximal end of the radius and ulna, except for some taxa that present a single insertion at the proximal end of the ulna (e.g., Struthio, [18]). The main function of this muscle in extant archosaurs is to flex the forearm and, in turn, contributes to the protraction of the arm [18, 20, 41, 43], Table 1.
It is inferred that in Bambiraptor, Buitreraptor, Archaeopteryx, and Confuciusornis the origin of the m. biceps brachii was at the end of the acrocoracoid process (Fig. 2), in agreement with the original proposals of Jasinoski et al. [18], Nicholls & Russell [23] and Ostrom [1].
In Buitreraptor, there are no muscular scars referring to the m. biceps brachii on either the ulna or radius, therefore the insertion site of this muscle cannot be established with certainty [56]. Moreover, Bambiraptor, Archaeopteryx and Confuciusornis have a tubercle at the proximal end of the radius that would indicate the insertion of the m. biceps brachii [50, 55, 63]. Considering this characteristic and based on extrapolation from extant archosaurs, it is highly probable that in paravians and basal birds this muscle is anchored at the proximal end of the radius, although insertion into the ulna would be doubtful (see [37, 50, 63]).
The function of the m. biceps brachii in Bambiraptor, Buitreraptor, Archaeopteryx and Confuciusornis probably was to flex the forearm, protract, and depress the humerus (Table 1). This is due to the cranioventral position of the end of the acrocoracoid process with respect to the glenoid cavity, consistent with the function proposed by Ostrom [1], Nicholls & Russell [23] and Jasinoski et al. [18].
Our results suggest that the anchorage of pectoral muscles were remarkably similar in the non-avian paravians Buitreraptor and Bambiraptor, and the basal avialan Archaeopteryx and Confuciusornis, being different from most Ornithothoraces (see also [10, 11, 20]).