
You have full access to this article via your institution.
Mouse embryonic stem cells maintain lineage immortality through sporadic entry into a two-cell-like state that enables asymmetric partitioning of DNA damage between daughter cells: one destined for elimination and one functionally rejuvenated.
A fundamental paradox in biology is how cell lineages achieve immortality while individual cells inevitably accumulate damage. Embryonic stem cells (ESCs) represent one of nature’s solutions to this problem: they can proliferate indefinitely in culture while maintaining genomic stability and developmental potential. The mechanisms underlying this remarkable property remain enigmatic, particularly given that ESCs are continuously exposed to replication stress and spontaneous DNA damage.
In this issue of Cell Research, Min, Zhang, and colleagues report an elegant resolution to this paradox.1 Using sophisticated long-term live-cell imaging combined with fluorescent reporters, they demonstrate that mouse ESCs periodically enter a transient two-cell-like (2C-like) state that serves as a critical quality-control checkpoint for the stem cell lineage.
The 2C-like state has long puzzled stem cell biologists. Marked by expression of genes like MERVL and Zscan4,2 this state occurs spontaneously in approximately 1% of cultured ESCs at any given time. Previous studies showed that blocking entry into this state triggers culture crisis,3 while cells in the 2C-like state paradoxically exhibit elevated DNA damage and increased apoptosis.4 These seemingly contradictory observations raised a fundamental question: how does a state characterized by high damage and death contribute to long-term stem cell renewal?
Through meticulous single-cell lineage tracking over 4-day movies, the authors discovered that 2C-like cells undergo functionally asymmetric divisions, producing two daughter lineages with dramatically different fates (Fig. 1). Approximately 60% of cells entering the 2C-like state divide asymmetrically, generating one daughter lineage (termed “2C-death”) that accumulates high levels of MERVL expression, exhibits extensive DNA damage, and ultimately undergoes cell death. The sister lineage (termed “2C-survived”) shows diminishing MERVL expression, reduced DNA damage, and returns to the pluripotent state with enhanced functional properties.
Fig. 1: Asymmetric division in the 2C-like state enables ESC lineage rejuvenation.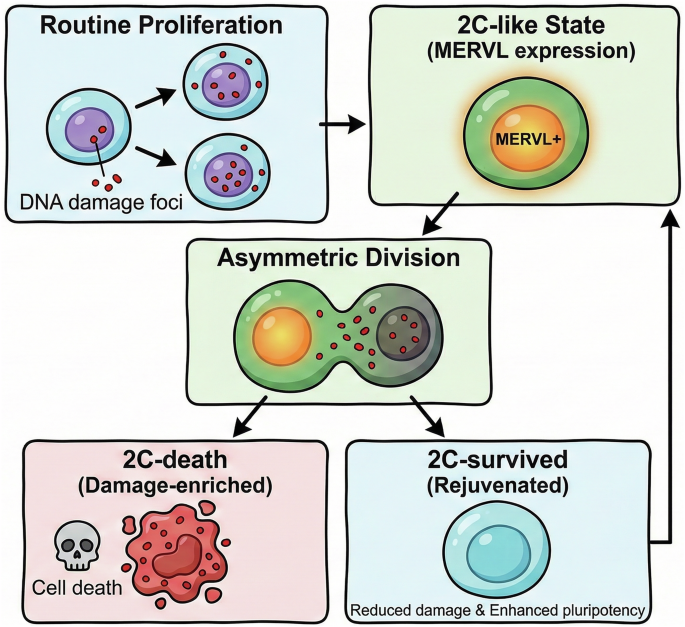
During routine proliferation, ESCs accumulate DNA damage (red foci). Upon entering the 2C-like state (marked by MERVL expression), cells undergo asymmetric division that segregates damage asymmetrically between daughters. One lineage becomes enriched for damage and undergoes cell death (2C-death), while the sister lineage (2C-survived) exhibits reduced damage and enhanced pluripotency, contributing to long-term lineage renewal.
The key mechanistic insight came from live-cell imaging of DNA damage using a 53BP1-mVenus reporter. The authors found that damaged DNA, visualized as 53BP1 foci, segregates asymmetrically during 2C-like cell divisions, with the majority of damage-containing foci preferentially inherited by the 2C-death lineage (Fig. 1). Remarkably, this asymmetric segregation requires an intact DNA damage response pathway: inhibiting ATM, ATR, CHEK, or PARP significantly reduced both the frequency of asymmetric divisions and the degree of damage asymmetry between sisters.
The functional consequences of this asymmetric division are striking. Using a clever fluorescent protein timer system that distinguishes cells based on how recently they exited the 2C-like state, the authors demonstrate that 2C-survived cells exhibit hallmarks of rejuvenation: reduced DNA damage by comet assay, enhanced alkaline phosphatase activity, elevated expression of pluripotency markers like Nanog and Oct4, improved clonogenicity in vitro, and most impressively, 73% chimeric efficiency when injected into blastocysts, compared to only 13% for cells that had not recently transited through the 2C-like state.
This work elegantly connects cellular aging mechanisms across phylogeny. Just as budding yeast segregates damaged proteins asymmetrically to generate rejuvenated daughters at the expense of aging mothers,5 ESCs leverage asymmetric division to maintain lineage youth despite individual cell aging. The similarity extends to the molecular machinery: the authors show that old centrosomes preferentially segregate to the 2C-death lineage, reminiscent of spindle pole body aging in yeast.
Several important questions emerge from this work. First, what molecular mechanisms drive the asymmetric partitioning of damaged DNA? The requirement for DNA damage response proteins suggests active sensing and sorting, but the downstream effectors remain unknown. Second, does asymmetric division during the 2C-like state represent a conserved rejuvenation mechanism in other stem cell types, or even in vivo during early development? Third, could the manipulation of this process, either enhancing asymmetric division to combat stem cell exhaustion6 or blocking it to target cancer stem cells, have therapeutic applications?
The quantitative framework provided by the authors is particularly compelling. They calculate that rejuvenation persists for 8–10 generations, meaning only 0.1%–0.4% of cells need to undergo rejuvenation per generation to maintain population health, well within the observed 1% 2C-like fraction. This demonstrates that sporadic entry into the 2C-like state is quantitatively sufficient to power long-term ESC self-renewal.
This study fundamentally reframes how we understand cellular immortality. Rather than relying solely on efficient damage repair, ESCs employ a division-based strategy that concentrates damage into disposable lineages while regenerating pristine ones. Importantly, these findings help clarify a previous confusing observation that “the Zscan4+ state is the process that restores potency, but the process itself (that is, Zscan4+ state) has no or low potency”7: cells in the 2C-like state indeed have compromised function, but asymmetric division ensures that one daughter (2C-survived) downregulates 2C marker expression and escapes with enhanced potency while the damaged sister (2C-death) retains high levels of 2C markers and is eliminated. As the first demonstration of functionally asymmetric division driving rejuvenation in mammalian cells, this work establishes ESCs as a powerful model for studying cellular aging and renewal, with potential implications for regenerative medicine and cancer biology.